Федяков.Шуктомов. Попытка применения показателя Престона для сравнения фаун и зоогеографических особенностей моллюсков и полихет некоторых районов западной части Белого моря. 1972.
Специализированная школа-интернат при Ленинградском
государственном университете
Биологический кружок
Попытка применения показателя Престона для
сравнения фаун и зоогеографических особенностей
моллюсков и полихет некоторых районов
западной части Белого моря
Ленинград 1972
Настоящая работа представляет собой попытку разобраться в закономерностях распределения фауны в Кандалакшском и Онежском заливах Белого моря.
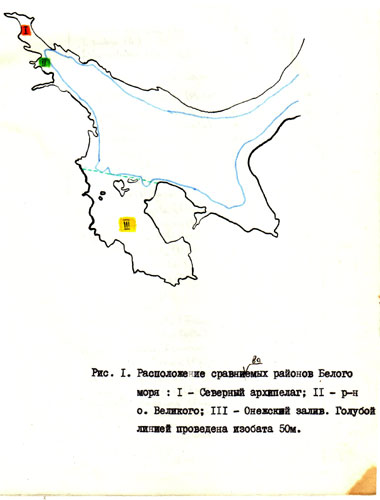
Зоогеографический состав фауны Белого моря очень неоднороден /Дерюгин, 1928; Гурьянова, I948/. Неоднородными в этом отношении оказываются не только Бассейн и Воронка Белого моря, но и его заливы. В кутовой части Кандалакшского залива было отмечено некоторое преобладание арктического элемента фауны и снижение численности и биоценотической роли бореальных видов по сравнению с более открытыми частями залива /Нинбург и др., в печати/. Для объяснения этого неожиданного для мелководного района факта была выдвинута следующая гипотеза. В куту Кандалакшского залива мог сохраниться арктический комплекс Йолдиевого моря, тогда как в Онежском сохранились в большей степени элементы бореальной фауны более позднего Литторинового моря. При этом, если миграции арктических видов и расселению их по всему району ничего не препятствовало, то для проникновения бореальных видов из Онежского залива вглубь Кандалакшского препятствовали низкие температуры больших глубин, разделяющих эти мелководные районы. Этот предполагаемый барьер хорошо виден на карте, на которую нанесена изобата 50 м, глубже которой преобладают отрицательные температуры /рис, I/.
Чтобы выяснить, действительно ли не происходит смешения различных фаун этого района или же существует единая для всего Кандалакшско-онежского района фауна, были выбраны две наиболее изученные в фаунистическом отношении группы - моллюски и полихеты. Мы сравнивали фауну в следующих районах: Северный архипелаг, район о.Великого и Онежский залив. При этом за границу Онежского залива мы приняли линию, проходящую через северную оконечность о. Анзерского. Сделано это было из тех соображений, что в противном случае в фауну залива окажутся включенными виды, приуроченные к арктическим водам Бассейна. Первоначально мы намеревались включить в анализ еще один район – район Гридинской губы /Русанова, 1949; Кобякова, 1949/, но сделать это не удалось из-за явно недостаточных данных /неполные списки видов/ по этому району.
Для оценки различий между фаунами отдельных районов был выбран коэффициент Престона /Старобогатов, 1970/, который находится по следующей формуле:
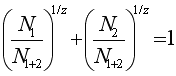
где N1- количество видов в одной районе, N2 - количество видов в другом районе, N1+2 - суммарное количество видов в двух районах /поскольку смысл проводимого Престоном сравнения состоит в сопоставлении отдельных фаун с фауной единой для сравниваемых районов, естественно, что виды, встреченные в обоих районах учитываются только один раз/. Показателем различия считается Z , причем если Z ≤ 0.27, районы считаются фаунистически однородными, тем более, чем ближе Z к 0. Если же Z > 0.27 фауны считаются изолированными друг от друга. Заметим, что выбранный показатель не зависит от величины исследованной площади, что делает его особенно удобным. Недостатком коэффициента Престона, очевидно, является произвольный выбор граничного значения = 0.27,
Необходимость проверять, одинаково или по разному протекают процессы обмена видами между фаунами для видов разного зоогеографического происхождения привела нас к мысли помимо общего анализа всей фауны проделать таковой для видов арктических и видов бореальных в отдельности.
В таблице I приводятся списки видов моллюсков и полихет для двух наиболее изученных районов Кандалакшского залива и для Онежского залива.
Виды |
Зоогеографическая х-ка |
Северный архипелаг |
Великий |
Онежский залив |
|
Mollusca: Tonicella mamorea (Fabr.) |
А-Б |
+ |
+ |
+ |
|
T. rubra (L) |
А-Б |
|
|
+ |
|
Lophirochiton albus (L) |
А-Б |
+ |
+ |
+ |
|
Chaetodema mitidium Loven |
А-Б |
|
+ |
+ |
|
Punctuella noaechina (Phipps) |
А-Б |
+ |
+ |
+ |
|
Margarites helicinus (Pipps) |
А-Б |
+ |
+ |
+ |
|
M. groenlandicus (Chemnitz) |
А-Б |
+ |
+ |
+ |
|
M olivaceus (Brown) |
А-Б |
+ |
+ |
+ |
|
M. cinereus (Couth) |
А-Б |
+ |
+ |
+ |
|
M. vahlii (Moller) |
А-Б |
? |
|
+ |
|
Solariella obscura (Cauth) |
А-Б |
|
+ |
+ |
|
S. varicosa (Migh. et Ad.) |
А |
? |
+ |
+ |
|
Ganesa laevigata (Friele) |
А-Б |
? |
|
+ |
|
Acmaea teshedinalis (Muller) |
А-Б |
+ |
+ |
+ |
|
Lepeta coeca (Muller) |
А-Б |
+ |
+ |
+ |
|
Molleria costulata (Muller) |
А-Б |
+ |
+ |
+ |
|
Littorina littorea (L) |
Б |
+ |
+ |
+ |
|
L. saxatilis (Maton) |
Б |
+ |
+ |
+ |
|
L. palliata (Say) |
Б |
? |
|
+ |
|
L. obtusata (L) |
Б |
+ |
+ |
+ |
|
Lacuna divaricata (Fabr.) |
Б |
+ |
+ |
+ |
|
L. pallidula (Da – Costa) |
Б |
+ |
+ |
+ |
|
Hydrobia ulvae (Pemaut) |
Б |
+ |
+ |
+ |
|
Homologira atomeus (Philippi) |
- |
+ |
+ |
+ |
|
Jeffresia globularis Jeffr. |
- |
+ |
+ |
+ |
|
Alvania jan-majeni (Friele) |
А |
+ |
+ |
+ |
|
Skeneopsis planorbis (Fabr.) |
Б |
+ |
+ |
+ |
|
Trichotropis borealis Brod et Sow |
А |
+ |
+ |
+ |
|
Tr. herzensteini Derjugin et Gurjanova |
А |
? |
|
+ |
|
Tr. kroyeri Philippi |
А |
? |
+ |
|
|
Natica clausa Brod. et Sow. |
А-Б |
+ |
+ |
+ |
|
Polynices pallidus (Brod. et Sow.) |
А-Б |
+ |
+ |
+ |
|
P. nanus (Muller) |
Б |
? |
|
+ |
|
Acribia islandica (Gmelin) |
А-Б |
+ |
+ |
+ |
|
Velutina velutina (Muller) |
А-Б |
+ |
+ |
+ |
|
V. nudata (Brown) |
А-Б |
+ |
+ |
+ |
|
V. plicatilis (Muller) |
А |
+ |
+ |
+ |
|
Trophonopsis clathratus (L) |
А |
+ |
+ |
+ |
|
Tr. truncatus (Strom) |
А-Б |
+ |
+ |
+ |
|
Columbella rosacea (Gould) |
А-Б |
+ |
|
+ |
|
Buccinum undatum (L.) |
Б |
+ |
+ |
+ |
|
B. groenlandicum Chemn. |
А |
+ |
+ |
+ |
|
B. glaciale (L.) |
А |
+ |
+ |
+ |
|
B. tenue Gray |
А |
+ |
+ |
+ |
|
Neptunea despecta (L.) |
Б |
+ |
+ |
+ |
|
Sipho latericeus (Moller) |
А-Б |
+ |
+ |
+ |
|
Achuete viridula (Fabr.) |
А |
+ |
+ |
+ |
|
Lora pyramidalis Strom |
- |
+ |
+ |
+ |
|
L. impressa Morech |
- |
|
|
+ |
|
L. trevelyana (Turton) |
- |
+ |
|
+ |
|
L. decussata (Couth.) |
- |
|
+ |
+ |
|
L. scalaris (Moller) |
- |
+ |
|
+ |
|
L. horpa Dall |
- |
|
|
|
|
L. exarata (Moller) |
- |
+ |
+ |
+ |
|
L. nobilis Muller |
- |
|
+ |
+ |
|
L. violacea (Mighels) |
- |
+ |
|
+ |
|
L. rugularia Fr. |
- |
|
|
+ |
|
L. tenuicostata (Sars) |
- |
+ |
|
+ |
|
Lora harpularia (Couth.) |
- |
+ |
+ |
+ |
|
Cylichua alba (Brown) |
А-Б |
+ |
+ |
+ |
|
C. scalpta (Reeve) |
А |
+ |
+ |
+ |
|
Retusa pertenuis (Mighels) |
А |
+ |
+ |
+ |
|
Diaphana hyalina (Turton) |
А-Б |
+ |
|
|
|
Philine fimuarchica G. Sars |
А-Б |
|
|
|
|
Ph. lima (Brown) |
А |
+ |
+ |
+ |
|
Scaphander lignaris L. |
Б |
+ |
+ |
+ |
|
Cadlina laevis (L.) |
А-Б |
+ |
+ |
+ |
|
Onelridoris muricata (Mull.) |
Б |
+ |
+ |
+ |
|
O. fusca (Muller) |
Б |
+ |
|
+ |
|
Palio dubia (M. Sars) |
Б |
|
+ |
+ |
|
Acantodoris pilosa (Mull.) |
Б |
+ |
+ |
+ |
|
Ancula eristata (Alder) |
Б |
+ |
+ |
+ |
|
Heolidia papillosa (L.) |
Б |
+ |
+ |
+ |
|
Coryphella rufibranchialis(Johsrst.) |
Б |
+ |
+ |
+ |
|
Coryphella borealis Odhner |
Б |
|
+ |
|
|
Dendronotus frondosus (Aseanius) |
Б |
+ |
+ |
|
|
Nucula tenuis (Montagu) |
А-Б |
+ |
+ |
+ |
|
Leda pernula (Muller) |
А-Б |
+ |
+ |
+ |
|
Portlandia arctica (Gray) |
А |
+ |
+ |
|
|
Joldia hyperborea (Loven) |
А |
+ |
+ |
+ |
|
Chlamys islandicus (Mull.) |
А-Б |
+ |
+ |
+ |
|
Anomia squamula (L.) |
А-Б |
+ |
+ |
+ |
|
Mytilus edulis (L.) |
А-Б |
+ |
+ |
+ |
|
Modiolus modiolus (L.) |
Б |
|
+ |
+ |
|
Musculus discors (L.) |
А-Б |
+ |
+ |
+ |
|
M. laevigatus (Gray) |
А-Б |
+ |
+ |
+ |
|
M. discrepaus (Gray) |
А-Б |
+ |
+ |
+ |
|
Crenella decussata (Montagu) |
А-Б |
+ |
+ |
+ |
|
Dacridium viltreum (Holboll) |
А-Б |
+ |
+ |
+ |
|
Cardium ciliatum Fabr. |
А |
+ |
+ |
+ |
|
Serripes groenlandicus (Chemm.) |
А |
+ |
+ |
+ |
|
Astarte crenata (Gray) |
А |
|
|
|
|
A. elliptica (Brown) |
А-Б |
+ |
+ |
+ |
|
A. montagui (Dillwyn) |
А |
+ |
+ |
+ |
|
A. borealis (Chemnitz) |
А |
+ |
+ |
+ |
|
Turtonia minuta (Fabr.) |
Б |
+ |
+ |
+ |
|
T. flexuosa (Montagu) |
К |
+ |
+ |
+ |
|
Macoma baltica (L.) |
Б |
+ |
+ |
+ |
|
M. calcarea (Chemn.) |
А-Б |
+ |
+ |
+ |
|
Mya arenaria (L.) |
Б |
+ |
+ |
+ |
|
M. truncata (L.) |
А-Б |
+ |
+ |
+ |
|
Saxicava arctica (L.) |
К |
+ |
+ |
+ |
|
Lyonsia arenosa (Moller) |
А |
+ |
+ |
+ |
|
L. schimkevitschi Derjugin et Gurjanova |
А |
|
|
|
|
Pandora glacialis (Leach.) |
А |
+ |
+ |
+ |
|
Cyprina islandica (L.) |
Б |
+ |
+ |
+ |
|
Polychaeta: Phyllodoce maculata L. |
Б |
+ ? |
+ |
+ |
|
Ph. groenlandica Oersted |
А-Б |
+ ? |
|
+ |
|
Ph. citrina Malmgren |
А-Б |
+ |
|
|
|
Ph. mucosa (Oersted) |
Б |
+ ? |
+ |
|
|
Eulalia bilineata Johnson |
Б |
+ |
|
+ |
|
Eu. viridis (L.) |
Б |
+ |
+ |
+ |
|
Eumida sanguinea (Oersted) |
Б |
+ |
+ |
|
|
Eteone longa (Fabricius) |
А-Б |
+ |
|
+ |
|
E. flava (Fabricius) |
А-Б |
|
+ |
+ |
|
E. barbata Malmgren |
А |
+ |
|
+ |
|
Antinoella badia Theel |
А |
+ |
+ |
|
|
A. sarsi Kinberg |
А-Б |
+ |
|
+ |
|
Eunoe nodosa Sars |
А-Б |
+ |
+ |
+ |
|
Gattyana cirrosa (Pallas) |
А-Б |
+ |
+ |
+ |
|
Lepidonotus squanuatus (L.) |
Б |
+ |
+ |
+ |
|
Harmothoe imbricata L. |
А-Б |
+ |
+ |
+ |
|
H. rarispina (Sars) |
А-Б |
+ |
|
+ |
|
Pholoe minuta Fabr. |
А-Б |
+ |
+ |
+ |
|
Glycera capitata Oersted |
К |
+ |
+ |
+ |
|
Pterosyllis formosa Clapharede |
Б |
+ |
|
|
|
P. finmarchica (Malmgren) |
А-Б |
|
|
+ |
|
Antolytus prismaticus (Fabr.) |
А-Б |
+ |
|
+ |
|
An. vercii Marenceller |
А |
|
|
+ |
|
Eusyllis blomstrandi Malmg. |
А-Б |
+ ? |
|
+ |
|
Sphaerosyllis erinaceus Claparede |
А-Б |
+ |
|
+ |
|
Castalia punctata Muller |
Б |
+ |
+ |
+ |
|
C. afroditoides Fabricius |
А-Б |
|
+ |
|
|
Nereis pelagica L. |
А |
+ |
+ |
+ |
|
N. virens Sars |
Б |
+ |
+ |
+ |
|
Nephthys longosetosa Oersted |
А-Б |
+ |
|
|
|
N. minuta Theel |
А |
+ |
+ |
+ |
|
N. caeca (Muller) |
А-Б |
+ ? |
|
+ |
|
N. ciliata (Muller) |
А-Б |
+ |
|
+ |
|
N. paradoxa Malmgren |
А-Б |
|
+ |
+ |
|
N. malmgreni Theel |
А-Б |
|
+ |
|
|
Ephesia gracilis Rathke |
А-Б |
+ |
+ |
|
|
Sphaerodorum minimum (Webst. et Bened.) |
А |
|
|
+ |
|
Lumbriconereis fragilis (Mull.) |
А-Б |
+ |
+ |
+ |
|
L. minuta Theel_ |
А |
+ |
|
|
|
Scoloplos armiger (Muller) |
К |
+ |
+ |
+ |
|
Nainereis guadricuspida (Fabr.) |
А-Б |
|
+ |
|
|
Polydora guadrilobata Jacobi |
А-Б |
+ |
+ |
+ |
|
Spio filicornis (Muller) |
А-Б |
+ |
+ |
+ |
|
Prionospio cirifera Wiren |
А-Б |
|
+ |
|
|
Pygospio elegans Claparede |
А-Б |
+ |
|
+ |
|
Microspio vireni (Augener) |
А |
|
|
+ |
|
Aricidea uschakovi Zachs |
А-Б |
+ |
|
+ |
|
Paraonis lyra Southern |
Б |
+? |
|
+ |
|
Diplocirrus longisetosus (Marenceller) |
А |
+ |
|
|
|
Flabelligera affinis Sars |
А-Б |
+ |
+ |
|
|
Brada granulate Malmg. |
А-Б |
+ |
|
+ |
|
B. villosa (Rathke) |
А-Б |
+ |
+ |
|
|
B. inhabilis (Rathke) |
А-Б |
|
+ |
|
|
Stylariaides plumosa (Mull.) |
А-Б |
+ |
+ |
+ |
|
Cossura longicirrata Webst. et Bened. |
А |
|
+ |
|
|
Cirratulus cirratus (Muller) |
А-Б |
+ |
+ |
+ |
|
Chaetozone setosa Malmg. |
А-Б |
+ |
+ |
+ |
|
Scalibregua inflatum Rathke |
А-Б |
|
+ |
+ |
|
S. robusta Zachs |
А |
|
|
+ |
|
Ophelia limacine (Rathke) |
А-Б |
+ |
+ |
+ |
|
Amnotrypaue aulogaster Rathke |
А |
|
|
+ |
|
Travisia forbesi Johnston |
А |
|
|
+ |
|
Capitella capitata (Fabr.) |
А-Б |
+ |
+ |
+ |
|
Heteromastus filiformis (Claparede) |
Б |
+ |
+ |
|
|
Notomastus lateciceus Sars |
К |
|
|
+ |
|
Nicomache lumbricalis (Fabr.) |
А-Б |
+ |
+ |
+ |
|
N. minor Arwidsson |
Б |
+ |
|
+ |
|
Praxillella praetermissa Malmg. |
А-Б |
+ |
+ |
+ |
|
P. gracilis Sars |
А-Б |
+? |
+ |
|
|
Rhodine gracilior (Tauber) |
Б |
|
+ |
+ |
|
Rh. loveni Malmgren |
Б |
+ |
+ |
|
|
Maldaue sarsi Malmgren |
А-Б |
+ |
+ |
+ |
|
Leiochone polaris (Theel) |
А |
|
+ |
+ |
|
Myriochele oculata Zachs |
А-Б |
+ |
+ |
+ |
|
Owenia fusiformis Delle Chiaga |
А-Б |
+ |
+ |
|
|
Pectinaria hyperborea Malmg. |
А-Б |
+ |
+ |
+ |
|
P. koreni (Malmgren) |
Б |
|
+ |
+ |
|
Anobothrus gracilis Malmg. |
А-Б |
+ |
|
+ |
|
Melinna elisabethae McIntosh |
А |
+ |
|
+ |
|
Ampharete acutifrous Grube |
А-Б |
+ |
+ |
+ |
|
A. linolstroemi Malmgren |
Б |
|
+ |
+ |
|
A. vega (Wiren) |
А |
|
|
+ |
|
A. arctica Malmgren |
А-Б |
|
|
+ |
|
Terebellides stroemi Sars |
А-Б |
+ |
+ |
+ |
|
Trichobranclius gracialis Malmg. |
А-Б |
+ |
+ |
|
|
Nicilea zostericola Oersted |
А-Б |
+ |
+ |
+ |
|
Pista maculata (Dalyell) |
А-Б |
+ |
+ |
|
|
P. flexuosa (Crube) |
А |
+ |
+ |
|
|
P. cristata (Muller) |
К |
|
|
+ |
|
Proclea graffi (Langerhaus) |
А-Б |
+ |
+ |
+ |
|
Polycirrus medusa Grube |
А-Б |
+ |
+ |
+ |
|
Leaena abrancliata Malmg. |
А-Б |
+ |
|
+ |
|
Neoamphitrite figulius (Dahyell) |
Б |
+ |
+ |
+ |
|
N. affinis Malmgren |
А-Б |
|
+ |
|
|
Laphania boeckii Malmg. |
А-Б |
+ |
|
+ |
|
Amphitrita cirrata Muller |
А-Б |
+ |
+ |
+ |
|
Zaonice cirrata (Sars) |
А-Б |
+ |
+ |
+ |
|
Artacama proboscidea Malmg. |
А-Б |
|
+ |
|
|
Terebella hessei Annenkova |
Б |
+ |
|
|
|
Chone infundibuliformis Kroyer |
А-Б |
+ |
+ |
+ |
|
Ch. duneu Malmgren |
А-Б |
+ |
+ |
+ |
|
Euchone amalis (Kroyer) |
А |
+ |
+ |
+ |
|
Eu. papillosa (Sars) |
А |
+ |
+ |
|
|
Sabella fabricii Kroyer |
А-Б |
+ |
|
+ |
|
Laomone kroyeu Malmgren |
Б |
+ |
+ |
|
|
Potanilla reniformis (Leuckart) |
Б |
|
+ |
|
|
Dasychone infarcta Kroyer |
А |
+ |
|
|
Чтобы избежать возможных неясностей, следует сразу оговорить, что приведенные в табл. 1 списки видев неполные. Так, из списка видов полихет исключены представители семейства Serpulidae, а из моллюсков - улитки рода Cingula. Поскольку определение этих животных связано с большими затруднениями, во многих работах они определены лишь до рода, кроме того трудно поручиться за полноту списка в каком-либо из выбранных районов. Естественно было исключить их из сравнения.
Виды рода Lorа и виды Homologira atomus и Jeffresia globularis учитывались только при сравнении общего состава фаун, но не учитывались при сравнении отдельных зоогеографических групп видов, так как их зоогеографическое происхождение пока неясно.
Наконец, напомним, что при анализе фауны Онежского залива мы учитывали не все виды, приводимые в работах Л.А.Кудерского /1966/ и Г. Сластниковым /1957/ а только те, которые встречены в мелководной части залива /т.е. южнее северной оконечности острова Анзерский/.
Результаты зоогеографического анализа отдельных районов сведены в таблицы II и III.
Таблица II
Зоогеографический состав фауны моллюсков отдельных
районов Белого моря
|
Район |
Северный архипелаг |
о. Великий |
Онежский залив |
|||
|
Зooгeoграф. принадлежность |
Видов |
% |
Видов |
% |
Видов |
% |
|
Арктическо-бореальные |
36 |
50.0 |
34 |
45.9 |
35 |
46.7 |
|
Арктические |
19 |
26.4 |
20 |
27.0 |
17 |
22.7 |
|
Бореальные |
15 |
20.8 |
18 |
24.3 |
21 |
29.3 |
|
Космополиты |
2 |
2.8 |
2 |
2.8 |
I |
1.3 |
Таблица III
Зоогеографический состав фауны полихет отдельных
районов Белого моря
|
Район |
Сев. архипелаг |
о. Великий |
Онежский залив |
|||
|
Зоогеогр. принадлежность |
Видов |
% |
Видов |
|
Видов |
% |
|
Арктическо-бореальные |
44 |
64.7 |
44 |
62.9 |
51 |
66.2 |
|
Арктические |
10 |
14.7 |
9 |
12.9 |
11 |
14.3 |
|
Бореальные |
12 |
17.6 |
15. |
21.4 |
11 |
14.3 |
|
Космополиты |
2 |
3.0 |
2 |
2.8 |
4 |
5.2 |
Данные, приведенные в таблицах II и III не позволяют достаточно точно судить о наличии различий в выбранных районах и о характере этих различий. Видно, что бореальные моллюски составляют незначительную часть малакофауны Северного архипелага, в районе Великого их доля несколько возрастает, достигая максимума в Онежском заливе. Эта тенденция хорошо иллюстрируется диаграммой /рис. 2/. Можно отметить также совсем незначительное уменьшение в Онежском заливе доли арктических видов моллюсков, тогда как в обоих районах Кандалакшского залива она остается практически постоянной. Все это во всяком случае не противоречит выдвинутой предварительно гипотезе. При сравнении фауны полихет указанных районов никакой явной тенденции не наблюдается, можно лишь отметить, что соотношение арктического и бореального элемента фауны ближе в районах Северного архипелага и Онежского залива, район же о.Великого стоит несколько особняком.
Прежде чем делать выводы, следует отметить, что только что приведенное сравнение делалось "на глаз" и не дает оснований о существовании, а тем более о величине изоляции между районами. Мы попытались оценить эти различия с помощью показателя Престона /поскольку прямое вычисление из вышеприведенной формулы чрезвычайно затруднительно, мы брали их из таблицы Престона, воспроизведенной в книге В.И.Старобогатова /1970/.
Таблица IУ
Значения показателя Престона, полученные при сравнении
полного списка видов каждого из районов Белого моря
|
|
Mollusса |
Ро1усhaeta |
||
|
|
Сев. архип. |
о.Великий |
Сев. архип. |
о. Великий |
|
Сев. apxип. |
|
0.28523 |
|
0.39511 |
|
Онежский зал. |
0.27688 |
0.21618 |
0.40165 |
0.50257 |
Из данных таблицы IУ следует, что между районом Северного архипелага и районами о. Великого и Онежского залива имеется незначительная изоляция в отношении фауны моллюсков. Между Великим и Онежским заливом изоляции нет и сходство этих фаун столь велико, что можно считать, что в результате свободного обмена видами фауны моллюсков слились в единую. В то же время фауны полихет всех трех районов достаточно изолированы друг от друга. Такую ситуацию можно объяснить тем, что моллюски группа арктическая и для миграций большинства видов моллюсков не существует термического барьера /говоря об арктическом происхождении моллюсков, мы имеем в виду, конечно, моллюсков Белого моря/. Полихеты же группа бореального происхождения, поэтому гораздо чувствительнее, чем моллюски к низким температурам. Для них холодные воды псевдоабиссали Белого моря, подходящие близко к берегу по обе стороны от о. Великого, создают неодолимый барьер.
Разобраться в особенностях распределения видов в западной части Белого моря помогает анализ бореального и арктического элемента фаун по отдельности /табл. Y и YI/
Таблица Y
Значения показателя Престона, полученные при сравнении
арктических видов, встречающихся в отдельных районах
|
|
MoIIusca |
Polychaeta
|
||
|
|
Сев. архип. |
о. Великий |
Сев. архип. |
о. Великий |
|
Сев. архип. |
|
0.23304 |
|
0.26240 |
|
Онежск залив |
0.28977 |
0.23293 |
0.76043 |
0.62432 |
Таблица YI
Значения показателя Престона, полученные при сравнении
бореальных видов, встречающихся в отдельных районах
|
|
MoIIusca |
Polychaeta
|
||
|
|
Сев. архип. |
о. Великий |
Сев. архип. |
о. Великий |
|
Север. архип. |
|
0.40592 |
|
0.48441 |
|
Онежск залив |
0.23478 |
0.23683 |
0.61954 |
0.45478 |
При анализе арктической части фауны моллюсков, видно, что изоляция, причем незначительная, существует лишь между районом Северного архипелага и Онежским заливом. Ранее приведенные данные /табл. III, рис. 2/ заставляют считать, что здесь правильнее говорить не об изоляции, а о различии, которое возникает постепенно - от кута к горлу Кандалакшского залива уменьшается доля арктических видов моллюсков. Четких, однако, границ провести нельзя. Что касается арктических видов полихет, картина оказывается совершенно иной. Между районами Великого и архипелага нет существенных различий в фауне арктической части полихет, в то же время фауны обеих частей Кандалакшского залива в большой степени изолированы от фауны Онежского. Очевидно, существует какой-то барьер, мешающий свободному обмену видами у арктического происхождения полихет между Кандалакшским заливом и Онежским, но какова его природа - сказать трудно.
Анализ распределения бореальных видов показывает, что для моллюсков существует барьер между районами Северного архипелага и о. Великого, точнее до Северного архипелага доходят далеко не все виды, встречающиеся в Онежском заливе и у о. Великого. Бореальные полихеты оказываются изолированными во всех трех районах, причем степень изоляции довольно велика. Это хорошо можно объяснить тем, что теплолюбивые виды полихет с трудом преодолевают низкотемпературные барьеры к северу от Великого и к югу от Керети.
Мы не приводим анализа арктическо-бореальных и космополитических видов, поскольку они, обладая наиболее широким ареалом, являются эврибионтными в большинстве своем; по этой причине анализ их распределения не несет никакой практически информации.
Попробуем сделать некоторые выводы из всего сказанного.
Во-первых, большинство фактов свидетельствует в пользу справедливости гипотезы о существовании на протяжении Кандалакшского залива двух низкотемпературных барьеров, которые мешают проникновению видов бореального происхождения из Онежского залива в направлении к куту Кандалакшского. Один из них расположен у северной оконечности о. Великого, а второй протянулся примерно от Керетского архипелага до Гридинской губы. Перемещениям арктических видов они не мешают, что подтверждается отсутствием изоляции в фауне арктических моллюсков между всеми тремя районами и незначительной изоляцией между арктическими видами полихет.
Анализ списков видов /табл. I/ наводит на мысль, что бореальные элементы фауны Кандалакшского залива представляют собой обедненный вариант соответствующего элемента фауны Онежского залива. Особенно четко это видно на моллюсках. В то же время судить об этом по показателю Престона нельзя, ибо он является ненаправленной величиной и не может указывать на возможные пути происхождения фаун.
Следует отметить, что к использованию любых численных показателей сходства /или различия/, в том числе и к показателю Престона надо относиться с осторожностью, ибо они в значительной степени формальны. По этой причине выводы настоящей работы нельзя считать окончательными.
В заключение мы хотим выразить благодарность Е. А. Нинбургу, оказавшему нам большую помощь при написании этой работы.

Литература
Броцкая В.А. Н.Н.Жданова, Н.Л.Семенова, 1963. Донная фауна Великой салмы и прилежащих районов Кандалакшского залива Белого моря. Тр. Кандад. гос. запов., в.IY; Тр. Беломорск. биол. ст. МГУ, т. II.
Зенкевич Л.А., 1956. Моря СССР, их фауна и флора. М.
Кудерский Л.А., 1966. Донная фауна Онежского залива Белого моря. Тр. Карельск. отд. ГосНИОРХ, т. IY, в. 2.
Нинбург Е.А., Гребельный С.Д., Биркан B.И., Иоффе Б.И.
Материалы к изучению донной фауны района Северного архипелага Кандалакшского залива Белого моря. В печ.
Сластников Г.С., 1957. Фауна Palychaeta Онежского залива Белого моря. Мат. по компл. изуч. Белого моря, вып. I.
Старобогатов Я.И., 1970. Фауна моллюсков и зоогеографическое районирование континентальных водоемов. Л.