Виноградов Г.М., Кобузева И.А., Сорокина О.С., Куприянова Е.А., Литвинова Е.Ю., Калякин М.В., Маколин О.И., Нигматов А.А. Современное состояние донных сообществ Ковдской губы Белого моря. 1. Каннское море и Горелая губа // В кн.: Состав и структура морского донного населения: сборник научных трудов (Ред. А.П.Кузнецов и О.Н.Зезина). 2001. М.: Изд-во ВНИРО. С. 110-117.
СОВРЕМЕННОЕ СОСТОЯНИЕ ДОННЫХ СООБЩЕСТВ КОВДСКОЙ ГУБЫ БЕЛОГО МОРЯ. 1. КАНСКОЕ МОРЕ И ГОРЕЛАЯ ГУБА.
Г.М.Виноградов[1], И.А.Кобузева[2], О.С.Сорокина2, Е.А.Куприянова[3], Е.Ю.Литвинова2, М.В.Калякин[4], О.И.Маколин2, А.А.Нигматов2.
Введение.
Ковдская губа расположена за Полярным кругом на Карельском берегу Кандалакшского залива Белого моря (рис. 1). Губа простирается в широтном направлении. Внешняя её часть, лежащая напротив места впадения в море реки Ковды и с. Ковда, разделена цепочкой островов на внутренний и внешний Ковдские рейды, а внутреннюю часть занимает большой о. Олений, отделенный от материка двумя неширокими проливами: Горелой губой на севере (глубина до 20 м) и Старцевой губой на юге. Горелая губа открывается в так называемое Канское море - небольшой внутренний бассейн глубиной до 12-15 м. Канское море ограничено о. Оленьим, мелкими островами кута Ковдской губы и материком. Между ним и собственно кутом Ковдской губы расположен мелководный Ягодный порог, на котором во время прилива и отлива возникает настоящая стремнина с бурунами. Небольшой порог существует и в Горелой губе недалеко от её окончания.
В начале XX века Ковдскую губу (включая Горелую губу и Канское море) подробно обследовала многолетняя экспедиция Юрьевского университета под руководством проф. К.К. Сент-Илера. Существующие описания этих работ [Сент-Илер, 1908, 1911, 1915], хотя и не дают полной картины донного населения губы в те годы, позволяют представить некоторые характерные его черты. Более подробное описание Горелой губы и Канского моря имеется в отчете студента Юрьевского университета А.И.Александрова, обширные выдержки из которого приводятся в опубликованном отзыве К.К. Сент-Илера [Сент-Илер, 1913]. Со времен этих работ в Ковдской губе произошли существенные гидрологические изменения, что могло отразиться на донном населении губы.
Самое существенное изменение касается солёности моря. До 1950-х гг. впадающая в губу р. Ковда очень сильно (до 0,2 - 6,3 о/оо) распресняла поверхностные воды Внутреннего Ковдского рейда, хотя в придонных слоях воды соленость приближалась к нормальной [Сент-Илер, 1934; Бухалова, 1949]. В 1950-х при строительстве Княжегубской ГЭС реку перекрыли плотиной, и большую часть воды направили в море другим путём (через канал в Княжой), что привело к заметному осолонению вод Ковдской губы (от 3 до 16, местами - до 21 о/оо в поверхностных слоях воды Внутреннего рейда, наши данные). Несмненно, изменилось и количество терригенного материала, выносимого рекой.
Однако на обстановке в Старцевой и Горелой губах, лежащих в стороне от основного потока воды из р. Ковды, перекрытие реки прямо отражаться не должно. Так, и в годы полноводной реки распресненные воды практически не попадали в Горелую губу [Сент-Илер, 1911] и понижение солености отмечалось только в самых поверхностных слоях воды в часы прилива (когда в губу входила распресненная вода с Ковдского рейда). Однако даже в поверхностных слоях Горелой губы соленость не спускалась ниже 9,2 о/оо [Бухалова, 1949], т.е., она никогда не была меньше критической величины 5-8 о/оо, при которой происходят заметные изменения в составе фауны [Хлебович, 1971]. Но в Горелой губе произошли свои изменения. В конце 1970-х г.г. о. Олений был соединен с материком практически сплошной дамбой (для протока воды оставлена единственная труба диаметром ~ 1 м). Дамба, разделившая собственно Горелую губу и Канское море, прошла по мелководному порогу Бродки (см. рис. 1). И, если до строительства дамбы приливное течение проходило Горелую губу и Канское море насквозь, то теперь оно входит и в Горелую губу, и в Канское море (вокруг о. Оленьего) как в “глухие” заливы с замкнутым кутом. Сплошной проток воды через эти участки моря прекратился, что, вероятно, должно было сказаться на донном населении губы.
В течении последних лет обширный бентосный материал, позволяющий оценить сегодняшнее состояние донных биоценозов и попытаться выявить возможные измсенения в их структуре, произошедшие со времен работ К.К. Сент-Илера, был собран в Ковдской губе во время летней практики специализированного биологического класса 520-й (а ранее - 57-й) школы г. Москвы. В настоящей работе рассматриваются биоценозы Горелой губы и Канского моря. Данные по фауне открытой части Ковдской губы, находящейся под влиянием р. Ковды, еще предстоит проанализировать.
Материал и методика.
Материал был собран во второй половине июля - августе 2000 г. Всего в Горелой губе и Канском море было получено 17 проб, из них 12 - ручной драгировкой с карбасов на глубинах 4 - 15 м, а остальные - на глубинах 0,5 - 1 м на Ягодном пороге и мелководьях Канского моря непосредственно со дна в часы отлива (рис. 1). Четыре пробы во внутренней части Горелой губы и три - в центральном районе Канского моря отбирали в тех же точках, что указаны в описаниях Сент-Илера [1908]. В пробах отмечался характер грунта, особое внимание уделялось наличию или отсутствию сероводорода. Животные определялись до вида и взвешивались. На основе этих данных высчитывались доли биомассы, приходящиеся на представителей основных экологических групп: эпибионтов (отдельно фильтраторов-сестонофагов, прикрепленных планктофагов, подвижных хищников, детритофагов, фитофагов) и инфауны илов и были построены спектры экологических групп, позволяющие наиболее наглядно представить структуру биоценозов [Свешников, Алигаджиев, 1986].
Кроме того, в различных частях губы были сделаны описания литорали и - в ясную безветренную погоду - проведены наблюдения с лодки ландшафа участков дна с глубиной в несколько метров.
Результаты.
Всего в собранной нами коллекции представлено 96 видов мезобентосных животных (табл. 1). Видовой состав, а так же спектры экологических групп мезобентоса Горелой губы и, особенно, Канского моря (рис. 2 а,б) типичны для илистых биотопов. Большая (до 26 % биомассы) доля хищников в Горелой губе (рис. 2а) определяется, в первую очередь, несколькими крупными Asterias rubens, попавшими в драги.
Донный осадок в районе работ (кроме Ягодного порога) был представлен преимущественно оливково-серым илом с незначительной примесью песка и гальки, в Горелой губе в пробах в очень большом количестве (более 1 кг на драгирование) присутствовали затонувшие щепки и куски коры - продукт деятельности лесозавода на о. Оленьем, существующего там с XIX века. Пробы, взятые в Горелой губе и центральном (лежащем напротив порога) районе Канского моря не имели запаха сероводорода. Очевидно, и при отсутствии прямого протока приходящей сюда в прилив свежей воды (в Горелую губу - со стороны фарватера и в Канское море - через Ягодный порог) оказывается достаточно для предотвращения развития сероводородного заражения. Однако некоторые признаки стагнации отмечены в окраинных частях этих акваторий. Так, осадок из окраинных районов Канского моря (в том числе из-под дернин зостеры и скоплений Cladophora) имел запах H2S, а на нижней литорали и верхней сублиторали здесь были отмечены небольшие пятна бактериальных матов. На илисто-каменистой литорали в куту Горелой губы так же встречались участки грунта с сероводородом. На небольшом пороге в Горелой губе на подходах к дамбе в часы прилива и отлива еще заметно сильное течение, и осадок здесь (в отличии от остальной губы) промыт, но перед самой дамбой на бывших Бродках начинает развиваться заиление. Здесь отмечен слой ила толщиной около 1 см, под котором располагается плотный песок. Наиболее заилёнными участками оказываются «карманы», примыкающие к Бродкам по бокам - сюда за счёт завихрения входного приливного течения наносит больше всего ила. На развитие стагнации указывает и то, что в кутовой части Горелой губы и в прилегающих к дамбе частях Канского моря отдельные участки дна, а так же зостера и фукусы часто, как ватой, покрыты мощными комьями нитчатых водорослей Cladophora fracta. Местами в выбросах по берегам Горелой губы сухая C. fracta образует широкие полосы, а в Канском море на поверхности плавают куски сбившегося из неё “войлока”, размером в первые десятки сантиметров, поддерживаемые на поверхности воды газовыми пузырями.
В устьевой части Горелой губы отмечены заросли ламинарий, здесь же было обловлено скопление Asterias rubens.
Дно Канского моря (на доступных для наблюдения с лодки глубинах) ровное, местами - с “полянками” Chorda filum, покрытое многочисленными створками раковин Macoma baltica, иногда - со скоплениями Asterias rubens. При приближении к входу в Ягодный порог расположен участок песчаного дна (тут дважды видели маленьких камбал). На мелководьях (особенно в небольших губах и проливчиках между островками кутовой части) обширные участки покрыты зарослями Zostera marina, причем в северо-западном куту Канского моря на зостере сидят многочисленные гидроиды Obelia geniculata. Среди зостеры регулярно попадаются немногочисленные мальки трёхиглой колюшки Gasterosteus aculeatus, а на листьях и стеблях зостеры прикрепляется молодь Mytilis edulus. Прибрежный пояс фукоидов развит довольно слабо. Ламинарий в Канском море нами отмечено не было, хотя их мощные заросли существуют по другую сторону ограничивающего его Ягодного порога. Присутствует Desmarestia aculeata. Из багрянок отмечена Polysiphonia arctica, а на участке, примыкающем к Ягодному порогу, где заметно течение - Phyllophora brodiaei, Polysyphonia urceolata и P. nigrescens.
Ягодный порог, через который Канское море соединяется с кутовой частью Ковдской губы, заслуживает отдельного описания. Он интересен прежде всего тем, что это редкий случай сильного течения морской воды на мелководье. На протяжении отлива течение идет из Канского моря. При этом слышен рёв воды на перекате, усиливающийся при приближении минимума отлива, а у камней возникают белые буруны. Немалую силу имеют и боковые завихрения течения, направленные в обратном к основному течению направлении. За несколько минут до минимума воды течение начинает замедляться, затем вода в стрежневой части пролива останавливается, и тут же начинается обратное перемещение воды - в сторону Канского моря. Уже через 10-15 мин сила течения заметно нарастает и буруны появляются вновь. При малой воде ширина порога сокращается в 3-4 раза, а отдельные валуны обсыхают. Порог покрыт водорослями Enteromorpha compressa, E. intectinalis, Monostroma fuscum (Chlorophyta), Fucus vesiculosus, F. inflatus, Chorda tomentosa, Pylaiella litoralis, Phyllitis fastia, Ascophilum nodosum (Phaeophyta), Rhodophilis dichotoma, Ahupheltia plicata, Polysyphonia urceolata (Rhodophyta). Отметим, что багрянки здесь проникают на очень малые глубины, а два первых вида не встречены в других местах кута Ковдской губы. Литораль, как и поверхность камней на самом пороге, почти сплошь укрыта щётками мидий, и весь порог представляет собой одну большую мидиевую банку. На камнях - очень мощные поселения Balanus balanoides в несколько слоёв. Среди мидиевых щеток встречены и вытянутые кривые Balanus crenatus. Огромное скопление мидий приводит к тому, что в спектре экологических групп обитающих здесь беспозвоночных абсолютно доминируют фильтраторы (рис. 2в). Другие встреченные здесь животные перечислены в табл. 1. Кроме беспозвоночных был обнаружен маленький пинагор Cyclopterus lumpus.
На подступах к порогу со стороны, противоположной Канскому морю, дно поросло ламинарией Laminaria saccharina (в основном) и L. digitata, которая отмечена только возле самой стремнины. В других местах внутренней части Ковдской губы L. digitata не встречается и присутствует только у “внешних” островков Ковдского рейда, там, где губа переходит в Кандалакшский залив ([Ковальчук, 1995] и наши данные). Мелководный стрежень, поросший ламинарией, тянется в сторону кутовой части Ковдской губы не менее чем на 300-400 м. В этом месте сохраняется сильный проток воды и, так же, как на самом пороге, обитают преимущественно животные-фильтраторы (рис. 2г). Но, в отличие от переката, здесь эта экологическая группа представлена в первую очередь не мидиями, а асцидиями и губками (табл. 1). Однако и мидии не исчезают полностью. Кроме Mytilis edulus здесь присутствуют Musculus laevigatus и Modiolus modiolus (редко встречающийся в Ковдской губе).
Ягодный порог оказывается неким “оазисом” со значительно большей биомассой донных беспозвоночных, чем окрестные воды. В таком качестве он оказывается весьма привлекательным для водоплавающих птиц - вблизи порога в период работ постоянно держалось (не проникая, однако, на участки с сильным течением) около 130 обыкновенных гаг Somateria mollissima и от полусотни до сотни турпанов Melanitta fusca, отмечались и другие утки. В этом же районе наблюдалось скопление (до 26 экз.) куликов-сорок Haematopus ostralegulus.
Обсуждение.
Сравнение наших данных (табл. 1) с описаниями К.К. Сент-Илера [1908, 1911, 1913] показывает, что целый ряд видов, отмеченных в Горелой губе в начале XX века, нами встречен не был. Конечно, наш материал далеко не столь обширен, как бывший в распоряжении К.К. Сент-Илера и его сотрудников, однако в нем нет и некоторых видов, которые были в Горелой губе обычными. Вероятно, они и в самом деле исчезли из губы, либо их численность здесь сильно снизилась. Это в первую очередь мшанки, брахиоподы Rhynchonella, гидроиды Campanularia, альционарии, актинии (исчезнувшие во всей Ковдской губе), Ophiura robusta (нами найдена только в устье Горелой губы) и Ophiocantha bidentata, Chlamys islandicus, Chelyosoma macleyanum, а так же Crossaster, Strongylocentrotus droebachiensis и ряд других видов; из иловых форм - Chirodota, Henricia, Urasterias lincki. Также не были найдены нами и обширные заросли багрянок, о которых сообщал А.И.Александров [цит. в: Сент-Илер, 1913] и Сент-Илер [1915], хотя отдельные экземпляры трех видов (Polisyphonia urceolata, Phyllophora brodiaei и Rhodophilis dichotoma), растущие на кусках коры, в пробах присутствовали. Нельзя, однако, исключить вероятность того, что в каких-то, оставшихся необследованными, участках Горелой губы заросли багрянок сохранились.
Можно заметить, что большинство ненайденных нами животных относится к экологическим группам, обычно приуроченным к местам с проточной водой, и, возможно, на их исчезновение повлияло прекращение прямого протока воды через Горелую губу. Возможно так же, что неблагоприятные условия в первую очередь сказались на зарослях багрянок, а угнетение ассоциированных с ними животных было вторично. Другое возможное объяснение - негативное влияние щепок и кусков древесной коры, в массе (более 1 кг на лов драгой) оказавшихся на дне губы в результате работы лесозавода на о. Оленьем. Щепки в донных пробах присутствовали здесь и во времена работ Юрьевского университета [Сент-Илер, 1908], однако за прошедшие годы их, вероятно, стало намного больше. Известно, что дампинг грунта, который в чем-то может быть аналогичен засорению поверхности дна остатками древесины, сильнее воздействует на эпибионтов, чем на роющих представителей инфауны [Оленин, 1989], а не встреченные нами животные принадлежат по большей части именно к первой экологической группе. Остаки древесины на дне могут негативно воздействовать и на таких обитателей илов, как Chirodota. С другой стороны, на кусках затонувшей древесины поселились изоподы-древоточцы Limnoria lignorum.
Таким образом, в Горелой губе намечается некоторая тенденция сдвига структуры биоценоза в сторону илистых форм, причем чем дальше от устья губы, тем сильнее. Канское море и в начале XX века было всё заселено типичной иловой фауной [Сент-Илер, 1913]. Ныне, когда оно, полностью отрезанно от былого протока воды через Горелую губу, эти его черты могут только усиливаться. И только Ягодный порог с его своеобразными условиями остался, судя по всему, неизменным и выполняет роль рефугия для ряда видов животных и водорослей, оставаясь единственным местом их обитания в кутовой части Ковдской губы.
Порог Бродки, по которому была построена дамба через Горелую губу, по описанию К.К. Сент-Илера [1915], весьма походил на Ягодный, но был уже и мельче его. Любопытно, что, при общей тенденции подъема берегов северо-западной части Белого моря (что, как отмечает Сент-Илер [1915], уже привело к полному или почти полному отделению от моря и последующему опреснению некоторых заливов Ковдской губы), через несколько десятков лет сообщение между Горелой губой и Канским морем могло бы прерваться естественным путем [Сент-Илер, 1915], но строительство дамбы резко ускорило этот процесс. Известна и достаточно хорошо изучена аналогичная ситуация в Долгой губе Соловецких островов, перекрытой дамбой в 1856 г. [Нинбург, 1990, 1997]. В частности, там тоже происходит выпадение некоторых видов, бывших (по данным Н.М.Книповича [1893]) весьма обильными 100 лет назад [Нинбург, 1997].
Вплоть до середины XX века Канское море было одним из наиболее рыбных мест Ковдской губы. Здесь в изобилии ловились камбала, треска, сельдь, навага, сиг, попадались корюшка, кумжа, зубатка, в массе обитали колюшки [Сент-Илер, 1908, 1915, 1934]. Жители Ковды часто ловили в нем рыбу. Отдельные рыбаки выходят в Канское море и сейчас, но, по их словам, рыбы в нем стало мало, и только вблизи Ягодного порога ловится навага. Меньше стало рыбы (в т.ч. трески) и в самой Ковдской губе. Эта картина вполне укладывается в существующую общую тенденцию оскуднения рыбных запасов северо-запдной части Белого моря [Лепская и др., 1995; Д.Лайус, Ю.Лайус, 1999], однако местное население однозначно связывает уменьшение количества рыбы с дамбой, перегородившей Горелую губу. Возможно, дамба действительно оказывает отрицательное влияние местного масштаба, например, влияя на кормовую базу рыб или не пропуская через Горелую губу из открытого моря рыб, которые могли бы размножаться и/или подрастать в относительно замкнутом и спокойном Канском море. Последнее предположение основано на том, что в Белом море прибрежно-островные районы с той или иной степенью гидродинамической замкнутости часто служат местом размножения рыб и важным источником их личинок для окружающих акваторий [Бояринцева, 1998], а о размножении “некоторых” рыб в Канском море вскользь упоминал К.К. Сент-Илер [1915]. Соотвествуют ли эти представления действительности - вопрос, несомненно требующий дальнейшего изучения.
Авторы благодарят Г.А.Соколову за неоценимую помощь при организации и проведении работ.
Список использованной литературы.
Бояринцева Е.А. Особенности удержания и выноса ихтиопланктона в прибрежно-островных районах Белого моря // Матер. юбилейн. научн. конф., посв. 60-летию Беломорской биол. станции им. Н.А.Перцова МГУ им. М.В.Ломоносова. - М.: Изд. ББС МГУ, 1998. - С. 18-20.
Бухалова В.И. Влияние опреснения на зоопланктон Ковдского залива Белого моря // Известия АН СССР: сер. биол. - 1949. - № 2. - С. 218-223.
Книпович Н.М. Несколько слов относительно фауны Долгой губы Соловецких островов и физико-географических её условий // Вестн. естествознан. - 1893. - Вып. 1-2. - С. 45-47.
Ковальчук Н.А. Описание ненарушенных промыслом ламинариевых сообществ, произрастающих у о. Микков (губа Ковда, Кандалакшский залив) // Проблемы изучения, рационального использования и охраны природных ресурсов Белого моря (материалы докладов). - СПб.: ЗИН РАН, 1995. - С. 123-124.
Лайус Д.Л., Лайус Ю.А. Что думают рыбаки о рыбе и рыбалке // Матер. IV научн. конф. Беломорской биол. станции МГУ 11-12 августа 1999 года. - М.: Русский Университет (Изд. ББС МГУ), 1999. - С. 98-101.
Лепская В.А., Зеленков В.М., Стасенков В.А. Современное состояние запасов промысловых рыб Белого моря // Проблемы изучения, рационального использования и охраны природных ресурсов Белого моря (материалы докладов). - СПб.: ЗИН РАН, 1995. - С. 133.
Нинбург Е.А. Долгая губа: изоляция естественная и искуственная // Природа. - 1990. - № 7. - С. 44-49.
Нинбург Е.А. Бентосное население Долгой губы и условия его существования // Матер. II научн. конф. Беломорской биол. станции им. Н.А. Перцова МГУ им. М.В.Ломоносова, 10-11 августа 1997 г. - М.: Изд. ББС МГУ, 1997. - С. 52-54.
Оленин С.Н. Применение морфоэкологического метода для оценки влияния сброса грунта на донную фауну Балтийского моря // Докл. АН СССР. - 1989. - Т. 306. - № 5. - С. 1276-1279.
Свешников В.А., Алигаджиев М.М. Жизненные формы беспозвоночных мезобентоса Каспийского моря // Докл. АН СССР. - 1986. - Т. 286. - № 2. - С. 482-486.
Сент-Илер К.К. Отчет об экскурсии на Белое море студентов-естественников Юрьевского университета летом 1908 года. - Юрьев: Типография К.Маттисена, 1908. - 67 с. (Повторено в: Уч. зап. Имп. Юрьевского универ-та. - 1909. - Т. 17. - № 3. - С. 1-67).
Сент-Илер К.К. Отчет об экскурсии студентов-естественников Императорского Юрьевского университета на Белое море летом 1911-го года. - Юрьев: Типография К.Маттисена, 1911. - 24 с. (Повторено в: Уч. зап. Имп. Юрьевского универ-та. - 1912. - Т. 20. - № 2. - С. 1-24).
Сент-Илер К.К. Отзыв об “Отчете А.И. Александрова, командированного Физико-Математическим факультетом для занятий по зоологии на Белое море летом 1912 года” // Уч. зап. Имп. Юрьевского универ-та. - 1913. - Т. 21. - Вып. 10. - С. 1-13.
Сент-Илер К.К. План исследования Ковденского залива и его окрестностей // Уч. зап. Имп. Юрьевского универ-та. - 1915. - Т. 23. - № 3. - С. 1-21.
Сент-Илер К.К. О результатах Беломорской экспедиции кафедры зоологии Воронежского университета летом 1934 г. // Труды Воронежского государственного университета. - 1935. - Т. 8. - Вып. 3. - С. 6-26.
Хлебович В.В. Концепция критической солёности в зоологии // Автореф. дисс. … д.б.н. - Л.: ЗИН АН СССР, 1971. - 36 с.
Таблица 1. Донные беспозвоночные, встреченные в Горелой губе и Канском море в 2000 г. (без учета фауны литорали).
|
Вид |
Экол. группа |
Устье Горелой губы |
Кутовая часть Горелой губы |
Канское море - центральная часть |
Заросли зостеры на мелководьях Канского моря |
Ягодный порог и заросли ламинарий на стрежне течения |
|
Spongia |
Ф |
+ |
+ |
+ |
|
# * |
|
Hydrozoa |
|
|
|
|
|
|
|
Dynamena pumila |
П |
|
|
+ |
|
+* |
|
Obelia geniculata |
П |
|
|
|
# |
+* |
|
Obelia loveni |
П |
+ |
|
|
|
|
|
Scyphozoa |
|
|
|
|
|
|
|
Lucernaria quadricornis |
П |
+ |
|
|
|
+ |
|
Nematoda gen.sp. |
И |
|
|
|
+ |
|
|
Nemertini |
|
|
|
|
|
|
|
Lineus gesserensis |
Х |
+ |
+ |
+ |
+ |
+ |
|
gen.sp. |
Х |
|
+ |
+ |
|
+ |
|
Priapulida |
|
|
|
|
|
|
|
Halicriptus spinulosus |
И |
|
|
+ |
|
+ |
|
Priapulus caudatus |
И |
|
+ |
# |
+ |
|
|
Polychaeta |
|
|
|
|
|
|
|
Anaitides maculata |
Х |
+ |
|
|
|
+ |
|
Antinoella sarsi |
Х |
|
+ |
|
|
+ |
|
Eunoe nodosa |
Х |
|
|
|
|
+* |
|
Harmathoe imbricata |
Х |
+ |
+ |
+ |
|
+* |
|
Lepidonotus squamatus |
Х |
+ |
|
|
|
+* |
|
Nephtys minuta |
Х |
# |
|
+ |
|
|
|
Nereis pelagica |
Х |
|
|
|
|
+ |
|
Pholoe minuta |
Х |
|
|
|
|
+ |
|
Brada granulata |
И |
|
|
# |
|
|
|
Diplocirrus longisetosus |
И |
+ |
|
|
|
|
|
Flabelligera affinis |
И |
+ |
|
|
|
|
|
Glycera capitata |
И |
# |
|
+ |
|
+ |
|
Lumbriconereis fragilis |
И |
|
+ |
|
|
|
|
Neoamphitrite figulus |
Д |
|
|
|
|
+ |
|
Polycirrus medusa |
Д |
|
|
# |
|
+ |
|
Polydora quadrilobata |
И |
# |
|
# |
|
+ |
|
Pygospio elegans |
И |
|
|
+ |
|
|
|
Spirorbis sp. |
Ф |
+ |
|
|
|
|
|
Stylarioides plumosa |
И |
+ |
|
|
|
|
|
Terebellides stroemi |
И |
# |
+ |
# |
|
|
|
Oligochaeta gen.sp. |
И |
|
|
|
+ |
|
|
Gastropoda |
|
|
|
|
|
|
|
Boreotrophon truncatus |
Д |
+ |
+ |
+ |
|
|
|
Buccinum sp. |
Д |
+ |
|
|
|
|
|
Cadlina laevis (?) juv. |
Х |
|
|
|
|
+ |
|
Cryptonatica clausa |
Х |
|
|
+ |
|
|
|
Cylichna alba |
И |
|
+ |
+ |
|
|
|
Cylichna occulata |
И |
|
# |
|
|
+ |
|
Dendronotus frondosus |
Х |
|
|
|
|
+ |
|
Epheria vincta |
Р |
+ |
+ |
|
|
+* |
|
Hydrobia ulvae |
Д |
+ |
+ |
+ |
# |
+ |
|
Lacuna nerotoidea |
Р |
|
|
|
|
+ |
|
Littorina littorea |
Р |
+ |
+ |
+ |
|
+* |
|
Littorina obtusata |
Р |
+ |
+ |
+ |
+ |
+ |
|
Margarites olivaceus |
Р |
+ |
|
|
|
+ |
|
Oenopota aff. pyramidalis |
Д |
|
+ |
|
|
|
|
Onoba aculeus |
Д |
|
# |
+ |
|
+ |
|
Skeneopsis planorbis |
Р |
|
|
+ |
|
+ |
|
Testudinalia tessellata |
Д |
|
|
|
|
+ |
|
Bivalvia |
|
|
|
|
|
|
|
Ciliatocardium ciliatum |
И |
+ |
+ |
|
|
|
|
Crenella decussata |
И |
|
+ |
+ |
|
|
|
Elliptica elliptica |
И |
|
|
+ |
|
|
|
Hiatella arctica |
И |
+ |
+ |
|
|
+* |
|
Macoma baltica |
И |
# |
# |
# |
# |
+ |
|
Modiolus modiolus |
Ф |
+ |
|
|
|
+ |
|
Musculus discors |
Ф |
+ |
|
|
|
|
|
Musculus laevigatus |
Ф |
+ |
+ |
+ |
|
+ |
|
Mya truncata |
И |
+ |
+/# |
+ |
|
|
|
Mytilis edulis |
Ф |
|
+ |
+ |
+ |
# * |
|
Portlandia arctica |
И |
|
+/# |
+ |
|
|
|
Serripes groenlandicus |
И |
|
|
|
|
+ |
|
Thyasira gouldi |
И |
|
+ |
+ |
|
|
|
Tridonta borealis |
И |
|
+ |
# |
|
+ |
|
Cirripedia |
|
|
|
|
|
|
|
Balanus balanoides |
Ф |
|
|
|
|
# * |
|
Balanus crenatus |
Ф |
|
|
|
|
+* |
|
Cumacea |
|
|
|
|
|
|
|
Diastylis glabra |
И |
|
# |
# |
|
|
|
Diastylis rathkei |
И |
+ |
|
|
|
|
|
Diastylis scorpioides |
И |
+ |
+ |
+ |
|
|
|
Diastylis sulcata |
И |
|
+ |
|
|
|
|
Isopoda |
|
|
|
|
|
|
|
Limnoria lignorum |
Др |
|
+ |
|
|
|
|
Amphipoda: Gammaridea |
|
|
|
|
|
|
|
Amphithoe rubricata |
Д |
|
|
|
|
+ |
|
Anonyx nugax |
С |
+ |
+ |
|
|
|
|
Atylus carinatus |
Д |
|
# |
# |
|
+ |
|
Centromedon pumilus |
С |
+ |
|
|
|
|
|
Corophium bonelli |
Д |
|
|
|
|
# |
|
Gammarelus homari |
Д |
|
|
|
|
+ |
|
Orchomene sp. |
С |
|
|
+ |
|
|
|
Pleustes sp. |
Д |
|
|
|
|
+ |
|
Pontoporeia femorata |
Д |
|
|
+ |
|
+/# |
|
Weyprechtia pinguis |
Д |
|
+ |
|
|
|
|
Amphipoda: Caprellidea |
|
|
|
|
|
|
|
Caprella septentrionalis |
Х |
|
|
|
|
# |
|
Decapoda |
|
|
|
|
|
|
|
Crangon sp. |
Д |
+ |
|
|
|
|
|
Pandalus annulicornis |
Д |
|
|
|
|
+ |
|
Pantopoda |
|
|
|
|
|
|
|
Nymphon sp. |
Х |
+ |
|
|
|
+ |
|
Insecta: Diptera |
|
|
|
|
|
|
|
личинки Chironomidae |
И |
+ |
|
+ |
+ |
+ |
|
Bryozoa |
|
|
|
|
|
|
|
Callopora craticula |
Ф |
+ |
|
|
|
+ |
|
Electra crustulenta |
Ф |
|
|
|
|
+ |
|
Escharella sp. |
Ф |
|
|
+ |
|
+ |
|
Lichenopora sp. |
Ф |
+ |
|
|
|
+ |
|
Shizoporella auriculata |
Ф |
|
|
|
|
+ |
|
Echinodermata |
|
|
|
|
|
|
|
Asterias rubens |
Х |
# |
+ |
+/# |
|
+ |
|
Ophiura robusta |
Д |
+ |
|
+ |
|
|
|
Tunicata |
|
|
|
|
|
|
|
Ascidia gen.sp. |
Ф |
|
|
+ |
|
|
|
Boltenia echinata |
Ф |
|
|
+ |
|
|
|
Dendrodoa sp. |
Ф |
|
|
|
|
+ |
|
Molgula arctica |
Ф |
+ |
+ |
|
|
|
|
Molgula siphonalis |
Ф |
|
+ |
# |
|
+ |
|
Molgula sp. |
Ф |
+ |
+ |
+ |
|
# |
|
Styela rustica |
Ф |
|
+ |
+ |
|
+ |
Примечания: “+” - присутствует; “#” - массовая форма, “*” в последней колонке стоит у видов, обнаруженных в т.ч. на самом перекате Ягодного порога. Экологические группы: И - инфауна илов, Др - древоточцы, прочие - различные эпибионты, среди которых дополнительно выделяются Ф - фильтраторы, П - прикрепленные планктофаги, Х - подвижные хищники, Д - детритофаги, Р - фитофаги, С – падальщики-трупоеды (стервятники).
СПИСОК ИЛЛЮСТРАЦИЙ К СТАТЬЕ Г.М.ВИНОГРАДОВА и др. “СОВРЕМЕННОЕ СОСТОЯНИЕ ДОННЫХ СООБЩЕСТВ КОВДСКОЙ ГУБЫ БЕЛОГО МОРЯ…”
Рис.1. - Схема расположения мест отбора проб в 2000 г. (очертания берегов Ковдской губы без кутовой части взяты из статьи К.К. Сент-Илера [1908]).
Рис. 2. Спектры экологический групп мезобентоса исследованных районов (по сырой биомассе, в %%). А - центральная часть Горелой губы; Б - центральная часть Канского моря; В - Ягодный порог (перекат); Г - заросли ламинарий на стрежне течения у Ягодного порога. 1-5 - эпибионты (1- фильтраторы-сестонофаги; 2 - прикрепленные планктофаги; 3 - подвижные хищники; 4 - детритофаги; 5 - фитофаги); 6 - интрабионты илов.
Present state of the benthic communities of the Kovdskaya Guba Bay (the White Sea). 1. Kanskoe More Bay and Gorelaya Guba Strait.
G.M. Vinogradov, I.A. Kobuzeva, O.S. Sorokina, E.A. Kupriyanova, E.Yu. Litvinova, M.V. Kalyakin, O.I. Makolin, A.A. Nigmatov.
Abstract
The benthic communities of the internal part of the Kovdskaya Guba Bay of the Kandalaksha Bay (the White Sea,) including shallow marine tidal rapid (threshold) were studied. We made an attempt to compare the present state of the bottom communities with the available descriptions made in the beginning of XX century. We try to estimate the influence of the dam which closed the water flow through the Gorelaya Guba Strait on these communities. Another source of influence were the remains of sank wood resulting from the industrial activity of the local timber mill. Several animal species were common in the Gorelaya Guba Strait in the beginning of XX century (different Bryozoa, Rhynchonella, Campanularia, Alcionaria, Actiniaria, Ophiura robusta, Ophiocantha bidentata, Chlamys islandicus, Chelyosoma macleyanum and some other species). However, we did not observe these species during our studies. Therefore, the shift toward mud-burrowing life-forms seemed to start. The species typical to firm bottom and flowing water remained at shallow-water Yagodnyi threshold.
УДК 591.5
РЕЗЮМЕ.
Исследованы бентосные сообщества внутренней части Ковдской губы Кандалакшского залива Белого моря, включая мелководный морской порог. Предпринята попытка сравнить современное состояние донных сообществ с имеющимися описаниями начала XX века и оценить возможное влияние на них дамбы, перекрывшей сквозной ток воды через Горелую губу, и покрывающих дно остатков утонувшей древесины - результат деятельности местного лесозавода. Ряд животных, обычных в Голрелой губе в начале XX века (мшанки, Rhynchonella, Campanularia, альционарии, актинии, Ophiura robusta, Ophiocantha bidentata, Chlamys islandicus, Chelyosoma macleyanum и др.) вновь обнаружен не был, и в структуре донных сообществ намечается сдвиг в сторону илистых форм. Типичные обитатели твёрдых грунтов и проточной воды сохранились на мелководном Ягодном пороге.
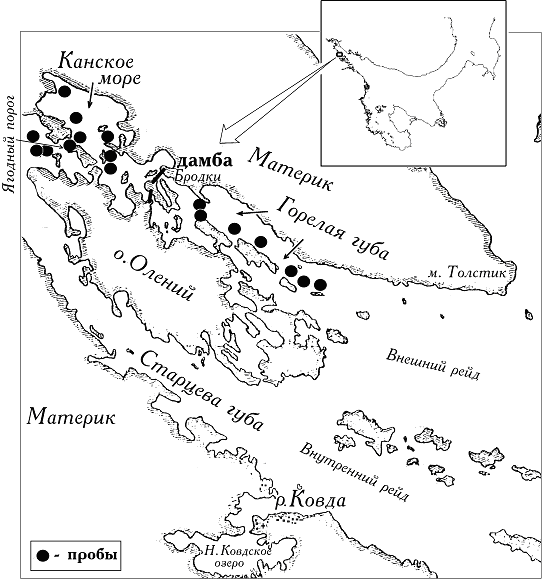
Рис.1 Схема расположения мест отбора проб в 2000 г. (очертания берегов Ковдской губы без кутовой части взяты из статьи К.К. Сент-Илера [1908]).
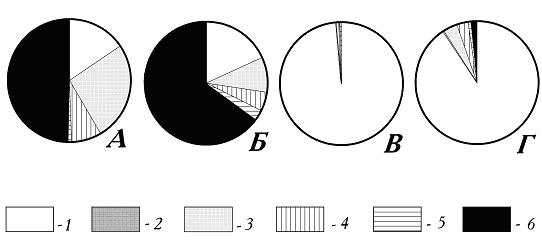
Рис.2 Спектры экологический групп мезобентоса исследованных районов (по сырой биомассе, в %%). А - центральная часть Горелой губы; Б - центральная часть Канского моря; В - Ягодный порог (перекат); Г - заросли ламинарий на стрежне течения у Ягодного порога. 1-5 - эпибионты (1- фильтраторы-сестонофаги; 2 - прикрепленные планктофаги; 3 - подвижные хищники; 4 - детритофаги; 5 - фитофаги); 6 - интрабионты илов.
[1] - Институт проблем экологии и эволюции им. А.Н.Северцова РАН;
[2] - Лицейский специализированный биологический класс московской школы № 520;
[3] - Российский Государственный медицинский университет;
[4] - Научно-исследовательский зоологический музей МГУ.