Алексеева Л. К вопросу о взаимосвязи Mytilus edulis и нитчатых водорослей. Санкт-Петербург. 2004.
Санкт-Петербургский Городской Дворец творчества юных
Лаборатория экологии морского бентоса
(гидробиологии)
Л. Алексеева
К вопросу о взаимосвязи Mytilus edulis и нитчатых
водорослей
Введение
Mytilus edulis L., 1758 - обычные обитатели прибрежной зоны, где они часто образуют массовые поселения. Прикрепляясь к какому-либо субстрату (камни, водоросли и т.д.) биссусами, они образуют так называемые щетки (друзы). Большие скопления мидий на литорали и на подводных мелководьях в более открытых частях моря носят название мидиевых банок (Зацепин, Филатов, 1968).
Замечательной особенностью поселений мидий является их аутоцикличность. Аутоцикличность - это самопроизвольное (без внешних воздействий) изменение размерно-возрастной структуры популяции во времени (Луканин, Наумов, Федяков, 1990).
Аутоцикличность развития мидиевых банок в Белом море хорошо изученная тема. Известно, что во времени происходит чередование четырех типов размерных структур мидий в банке. В зависимости от географических условий, течений, состава грунта и т. д. эти типы чередуются в разном порядке и с разной скоростью (Луканин, Наумов, Федяков, 1990). При этом одной из стадий цикла является такая размерная структура, в которой молодь мидий практически отсутствует. Отсутствие пополнения поселений молодью приводит к тому, что на банках периодически происходит вымирание старых генераций мидий (Луканин, Наумов, Федяков, 1990).
Авторы работ по аутоцикличности мидиевых поселений (Луканин, Наумов, Федяков, 1990) считают, что причина отсутствия молоди в поселении взрослых особей кроется в том, что старые моллюски выделяют репелленты, отпугивающие личинок.
Однако другими авторами было показано, что приток молоди в поселения, оформленные в виде друз, происходит в период разрастания нитчатых водорослей, которые предоставляют молодым мидиям место для оседания (Хайтов и др., 2002). А поскольку мидиевые друзы и мидиевые банки демонстрируют очень сходные закономерности динамики (Хайтов, 1999), то можно ожидать, что и в мидиевых банках будет наблюдаться аналогичная картина. В связи с этим становится крайне важным выявление механизмов связи нитчатых водорослей и поселений мидий.
Нитчатые водоросли - это группа водорослей, выделяемая условно, по сходству морфологического, реже - анатомического строения, а также по особенностям экологии и той роли, которую они играют в жизни донного населения (Голлербах, 1977). В группу "нитчаток", обитающих на Белом море, обычно включают представителей родов Cladophora, Acrosiphonia (зеленые водоросли), Ectocarpus, Dictyosiphon, Pylaiella, Stictyosiphon (бурые водоросли) и Polysiphonia (красные водоросли). Всего 16 видов водорослей (Возжинская, 1982).
Целью данной работы было выявление связи между цикличностью развития мидиевых банок и изменениями биомассы нитчатых водорослей. В рамках данной цели мы поставили следующие задачи:
1. описать многолетние изменения размерной структуры мидий на нескольких мидиевых банках;
2. описать изменения биомассы нитчатых водорослей на этих мидиевых банках;
3. проанализировать связь размерной структуры поселения мидий и обилия нитчатых водорослей.
Материал и методика
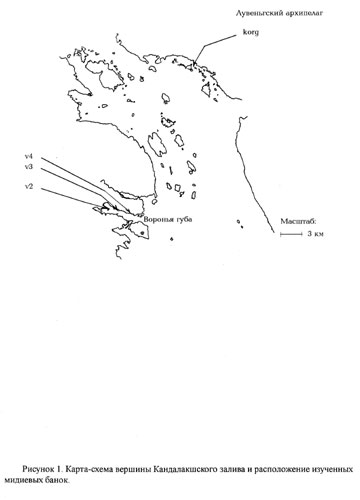
Материалом для данной работы послужили сборы, проведенные Лабораторией Экологии Морского Бентоса (гидробиологии) в 1997-2004 гг. Сбор материала проводился на нижней литорали островов и материка вершины Кандалакшского залива Белого моря на территории Кандалакшского природного заповедника (рис. 1). Всего было изучено четыре мидиевые банки.
Ежегодно на каждой банке бралось по 6 проб круглой рамкой с площадью захвата 1/182 кв. м. Точка взятия проб определялась случайно, путем бросания пробоотборника. Пробы брались только в тех местах, где наблюдался сплошной слой мидий. Если рамка падала на участок дна, не занятый мидиями, то проба бралась на ближайшем участке, покрытом слоем моллюсков. Рамка вдавливалась в грунт на глубину 10-15 см.
Грунт, попавший в рамку, промывался через сито с диаметром ячеи 0,5 мм. После промывки из проб, кроме мидий, выбирались все животные и нитчатые водоросли. После этого у мидий измерялась длина раковины с точностью до 1 мм. Нитчатые водоросли не сортировались по видам, была определена их суммарная биомасса. Водоросли высушивались на фильтровальной бумаге и взвешивались на торзионных весах с точностью до 1 мг. При большом количестве водорослей взвешивание производилось на аптечных весах с точностью до 10 мг.
Для описания размерной структуры мидий в банках в каждой банке объединялись все 6 проб, взятых в этой банке. Далее нами были построены гистограммы частотных распределений мидий на каждой станции за каждый год.
Для описания биомассы нитчатки мы использовали средние значения, вычисленные по шести пробам, на каждой банке за каждый год.
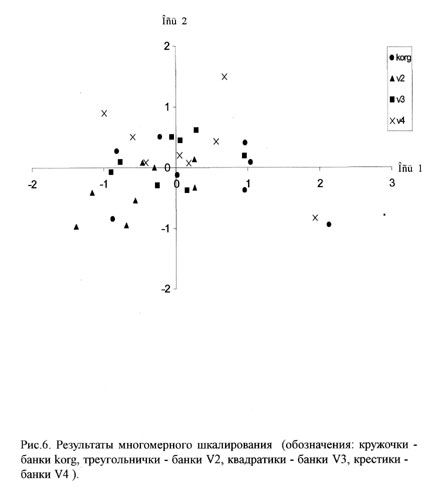
Для анализа варьирования размерных структур мидий мы использовали метод многомерного шкалирования. Многомерное шкалирование - это процедура, позволяющая наиболее эффективно разместить объекты на плоскости, приближенно сохраняя наблюдаемые между ними расстояния. Этот анализ был проведен нами с использованием пакета Statistica for Windows 6.0.
Чтобы получить изначальные данные для многомерного шкалирования, мы вычислили расстояния между объектами (мидиевыми банками), используя формулу евклидового расстояния:
Где f1i и f2i - это частота i-тых размерных классов на двух сравниваемых банках, выраженная в процентах.
Проведенное многомерное шкалирование позволило ранжировать все мидиевые банки в соответствии с их размерной структурой.
В заключение хотелось бы поблагодарить сотрудников Кандалакшского государственного природного заповедника за предоставление базы и помощь при проведении полевых работ. Также я благодарна всем участникам летних Беломорских экспедиций за первичную обработку материала. В особенности я признательна своему научному руководителю Вадиму Михайловичу Хайтову за неоценимую помощь при создании данной работы.
Изложение результатов
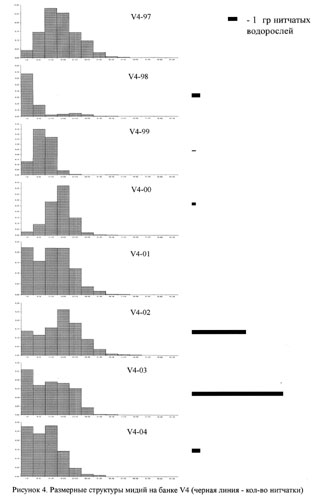
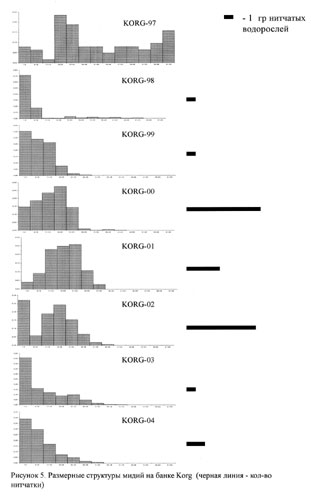
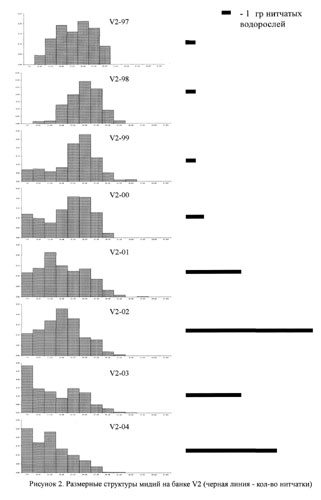
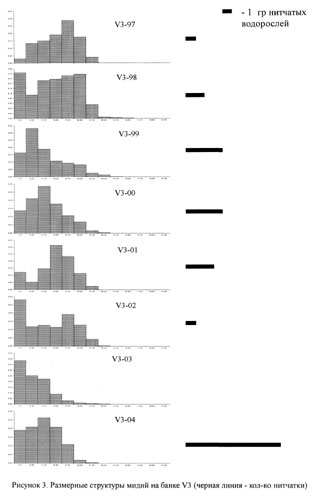
На рисунках 2-5 приведены размерные структуры мидий на разных банках в разные годы. Можно заметить, что размерная структура изменяется во времени: чередуются периодические вымирания отдельных совокупностей и периоды притока молоди.
На этом же рисунке приведены данные, которые позволяют сказать, что в разных банках есть периоды всплеска обилия нитчатых водорослей. В мидиевой банке V2 этот период приходится на 2001-2004 гг., в банке V3 - на 1998-2001 гг. и на 2004 год, в банке V4 - на 2002-2003 года и на банке Korg - 2000-2002 годах.
На рисунке 6 приведен график, отражающий многомерное шкалирование мидиевых банок в соответствии с размерной структурой мидий. Приведенные данные позволяют упорядочить все банки и присвоить каждой из них свой номер в соответствии с координатами по оси ОХ. Мы выбрали именно ось ОХ, т.к. разброс по оси OY значительно меньше и обеспечен отскакиванием описаний только лишь нескольких банок.
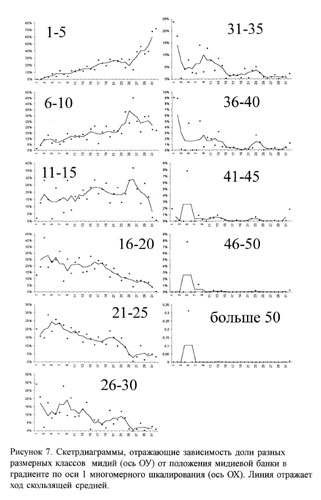
После описанного упорядочивания были построены скетр-диаграммы, на которых по оси абсцисс был отложен номер банки, а по оси ординат доля мидий тех или иных размерных классов (рис. 7). Можно заметить некоторые тенденции. Во-первых, по мере увеличения номера банки (увеличение координаты по оси 1 в многомерном шкалировании) увеличиваются доли молоди (1-5 и 6-10 мм) в мидиевом поселении. Во-вторых, уменьшаются доли мидий с размерами 16-20 и 21-25. Чуть хуже видно уменьшение доли мидий с размерами 26-30, 31-35, и 36-40. Так как доли мидий с размерными классами 50-55, 56-60 и 61-65 незначительны, мы построили общую скетрдиаграмму по их сумме (больше 50). Видно, что такие мидии встречаются в заметных количествах только в банках, занимающих левую часть градиента.
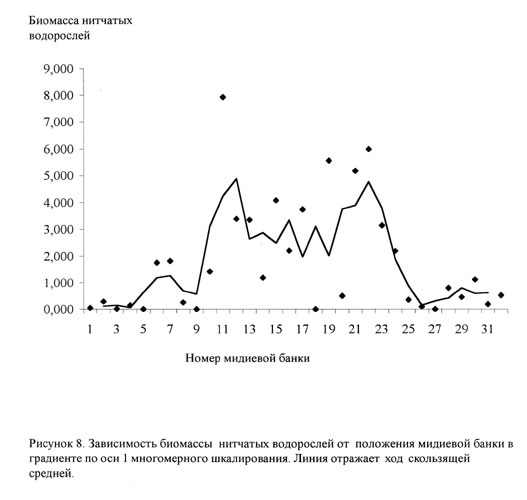
На скеттер-диаграмме, где по оси абсцисс был отложен номер банки, а по оси ординат количество нитчатых водорослей (рис.8). По этому графику можно выделить четыре совокупности. Первая, с небольшим количеством нитчатки - банки с номерами 1-10. Вторая и третья, с наибольшим количеством нитчатых водорослей -банки 11-17 и 18-24. Четвертая совокупность - банки, в которых нитчатки мало, с номерами 25-32.
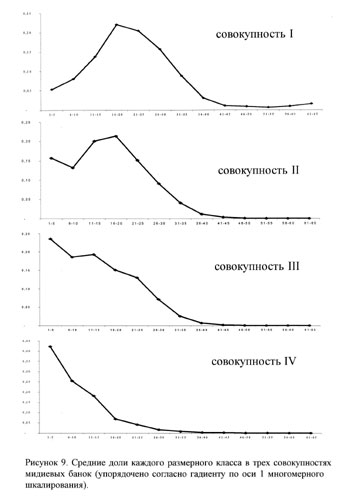
Для отражения размерной структуры, характерной для банок разных совокупностей, нами были выстроены четыре графика, отражающих средние доли мидий разных размерных классов для каждой совокупности банок (рис.9). Можно заметить, что для первой совокупности характерно обилие мидий с размерным классом 16-20. Для второй - обилие мидий с размерами 16-20 и немногим меньшее количество мидий с размерами 1-5. В третью совокупность попали банки с большим количеством мидий с размерами 1-5 и 11-15. А в четвертую - только те банки, в которых много молоди и очень мало других особей.
Распределение банок по совокупностям приведено в таблице 1.
Таблица 1. Распределение всех мидиевых банок в разные годы по совокупностям.
|
Мидиевые банки |
I совокупность |
II совокупность |
III совокупность |
IV совокупность |
|
Korg |
1997, |
2000, |
|
1998, |
|
|
2001. |
2002. |
|
1999, |
|
|
|
|
|
2003, |
|
|
|
|
|
2004 |
|
V2 |
1997, |
2001, |
2003, |
|
|
|
1998, |
2002, |
2004. |
|
|
|
1999, |
|
|
|
|
|
2000. |
|
|
|
|
V3 |
1997, |
1998, |
1999, |
2003. |
|
|
2001. |
2000. |
2002, |
|
|
|
|
|
2004. |
|
|
V4 |
1997, |
2002. |
2001, |
1998, |
|
|
2000. |
|
2003 |
1999, |
|
|
|
|
|
2004 |
Приведенные в этой таблице данные позволяют заметить, что все банки в начале наших наблюдений находились в первой совокупности, то есть они были сформированы в основном крупными мидиями.
Можно заметить, что в большинстве случаев после размерной структуры, соответствующей первой совокупности, формируется размерная структура, соответствующая второй или третьей совокупности. Наиболее четко эта закономерность видна на банке V2. Здесь размерная структура первой совокупности сменяется второй, а вторая третьей. Соответственно изменяется и обилие нитчатых водорослей.
Указанной закономерности противоречат лишь случаи на мидиевых банках Korg и V4, когда за размерной структурой, соответствующей первой совокупности, следовала размерная структура, соответствующая четвертой совокупности.
Обсуждение результатов
Как уже было описано в изложении результатов на всех изученных банках есть периоды всплеска обилия нитчатых водорослей. Эти всплески обилия нитчатки происходят в любой год, но не синхронно, а на разных банках индивидуально. Т.е. обилие нитчатых водорослей не зависит от каких-то внешних факторов, влияющих на банку, а, скорее всего, от локальных факторов в самой мидиевой банке.
Последовательность размерных структур согласуется с динамической последовательностью.
Известно, что рост растений зависит от концентрации азот- и фосфорсодержащих соединений, растворенных в воде (Peterson, Kenneth, 1999 цит. Хайтов и др., 2002). Значит, перед обильным ростом нитчатых водорослей в банке должны накапливаться в достаточном количестве азот и фосфор. Поскольку существует связь между обилием нитчатки и размерной структурой мидий, то можно предположить, что рост нитчатых водорослей приурочен к определенным стадиям цикла, описанного В. В. Луканиным и соавторами (Луканин, Наумов, Федяков, 1990). Полученные нами данные указывают на то, что безнитчатая стадия с размерной структурой I типа предшествует нитчатым (II и III типы) стадиям. Следовательно, именно на стадии, когда банка сформирована крупными мидиями, и происходит обогащение грунта азотом и фосфором, что «подготавливает» банку к разрастанию нитчатых водорослей.
После того, как нитчатка разрастается, на нее оседает молодь, формируя размерную структуру характерную для совокупностей II и III. А далее, по всей видимости, после разрастания нитчатых водорослей и оседания молоди, старые особи отмирают. Это приводит к тому, что на банке остаются только молодые моллюски (структура IV совокупности).
Таким образом, можно говорить о том, что теория аутоцикличного развития мидиевой банки (Луканин, Наумов, Федяков, 1990) может быть несколько дополнена. В соответствии с полученными нами данными цикл на мидиевой банке будет выглядеть следующим образом. На первой стадии в банке есть только крупные мидии, которые обогащают грунт азотом и фосфором. На второй стадии, на банке разрастаются нитчатые водоросли, на которые оседают молодые мидии. Далее старые моллюски отмирают, вместе с ними отмирает и нитчатка, а на банке остается только молодь. Далее молодые моллюски подрастают и вновь создают среду, благоприятную для роста нитчатых водорослей.
Выводы
1. Обилие нитчатых водорослей не зависит от каких-то внешних факторов, влияющих на банку, а, скорее всего, от локальных факторов в самой мидиевой банке.
2. На стадии, когда банка сформирована крупными мидиями, подготавливается среда для разрастания нитчатых водорослей. Вероятно, это связано с накоплением в грунте азото- и фосфорсодержащих соединений.
3. Разросшиеся нитчатые водоросли предоставляют субстрат для оседания молоди.
4. Теория аутоциклического развития мидиевой банки должна быть дополнена.
Список литературы
Голлербах ММ. Основные типы морфологической структуры тела водорослей. - Жизнь растений - том 3 - ред. Голлербах М. М. - М. , 1977 - стр. 32-37.
Зацепин В.И., Филатов ЗА. Класс Двустворчатые (Bivalvia) - Жизнь животных - том 2 - ред. Зенкевич Л.А. - М., 1986 - с. 95-155.
Луканин ВВ., Наумов АД., Федяков ВВ. Поселения мидий: постоянное непостоянство -Природа. - 1990, № 11 - с. 56-62.
Хайтов В.М., Артемьева А.В., Фокин М.В., Яковис ЕЛ. - Структурно-динамическая организация консорциев друз Mytilus edulis на литорали губы Долгой острова Большого Соловецкого. Ч / Структура и динамика поселений мидий -Вестник СПБГУ. Сер. 3.- 2002, вып. 4 (№ 27). - с. 9-20.
Хайтов В.М. - Сообщества донных беспозвоночных, связанные с естественными плотными поселениями мидий на мелководьях Белого моря (структура, динамика, биотические взаимоотношения). - Диссертация на соискание ученой степени кандидата биологических наук. - СПБ, 1999.
Peterson В. J., Kenneth L. H. Jr. The potential for suspension feeding bivalves to increase seagrass productivity // Journal of Experimental Marine Biology and Ecology. -1999.-№ 240.-p. 37-52
 |
 |
 |
| рис.1 | рис.2 | рис.3 |
|
рис.4 |
рис.5 |
рис.6 |
 |
 |
 |
| рис.7 | рис.8 | рис.9 |