Виноградов М.Е. Характер пищевых связей некоторых видов птиц с литоралью Белого моря // Труды Всесоюзного гидробиологического общества АН СССР, т. II, 1950. Стр.103-118.
М. Е. ВИНОГРАДОВ
ХАРАКТЕР ПИЩЕВЫХ СВЯЗЕЙ НЕКОТОРЫХ ВИДОВ ПТИЦ
С ЛИТОРАЛЬЮ БЕЛОГО МОРЯ
(Институт зоологии Московского государственного университета и Кандалакшский государственный заповедник)
Введение
Литораль — место «стыка» морской, наземной и воздушной, фаун. Благодаря постоянным или временным пищевым связям компонентов одной фауны с компонентами другой, этот «стык» не является пассивным соприкосновением. Напротив, он имеет характер системы сложных биологических взаимоотношений, связывающих эти фауны в единый биологический комплекс.
Настоящая работа представляет собой часть комплексной темы по изучению литорали Белого моря, проводимой кафедрой зоологии беспозвоночных МГУ и Кандалакшским гос. заповедником.
Работа преследует две цели: во-первых, осветить некоторые стороны биологии питания птиц и, во-вторых, определить значение потребляемой птицами пищи в общей экономике литорали. В соответствии с поставленными задачами определяется качественное использование птицами литоральной фауны, обосновывается разграничение всего орнитокомплекса на группы со сходными основными моментами биологии питания и дается количественное представление о степени выедания птицами литоральной фауны.
Работа проводилась летом (июнь — август) 1948 г. на Беломорской биологической станции МГУ, расположенной в Ругозерской губе Кандалакшского залива (66°34' с. ш. и 33° 10' в. д.). Были использованы также материалы, собранные П. В. Матекиным в 1946—1947 гг. в районе о-ва Лодейного в Кандалакшском заповеднике. Всего было просмотрено 211 желудков 20 видов птиц.
Берег Кандалакшского залива носит фиордовый характер (Гурьянова, 1948). Лес по берегам подходит к самой воде. Сильная изрезанность берега и тесный контакт леса с морем в значительной степени влияют на питание как литоральных, так и лесных птиц. Последние «прижимаются» к морю еще и вследствие того, что чернолесье, являющееся наиболее удобной стацией обитания для большинства лесных птиц, тянется узкой полосой в 100—1000 м вдоль берега; дальше, в глубь материка, идут болота, на которых птицы почти отсутствуют. Характер рельефа и распределение леса дают все градации связи различных птиц с литоралью и лесом.
Литораль, очень богато населенная различными организмами, является неистощимым источником питания и, естественно, при сравнительной бедности фауны насекомых в лесу, привлекает к себе лесных птиц. В то же время в прибрежных кустах часто наблюдаются массовые вылеты различных насекомых (пяденицы, огневки, мухи Bibio), делающихся легкой добычей морских птиц.
Своеобразный характер местности, где проводилась работа, в значительной степени определяет типы связей различных видов птиц с литоралью.
Биология питания птиц
Связь птиц с литоралью, очевидно, может осуществляться путем питания на литорали взрослых птиц и птенцов (у выводковых) и кормления птенцов литоральными животными (у птенцовых).
Сравнивая набор кормов у различных видов птиц, можно заметить, что он (особенно у птиц, наиболее тесно связанных с литоралью) отличается большим, как мы увидим дальше, однообразием. Это может быть объяснено лишь тем, что при огромных пищевых запасах литорали (277.3 г/м2 — по данным Абрикосова и Соколовой, 1948), связанным с ней птицам нет надобности узко специализироваться по определенным видам пищи, а наоборот, есть возможность всем в равной степени пользоваться чрезвычайно богатым набором кормов.
Но вместе с этим у всех видов птиц в пище можно выделить преобладание какого-либо одного вида или группы кормов, причем это преобладание столь значительно, что можно говорить об «основных» и «второстепенных» кормах.
Закономерность преобладания той или иной, пищи оказывается примерно одной и той же у многих видов, имеющих сходную биологию питания. У таких птиц частота встречаемости второстепенных для них кормов в общем тоже совпадает. Таким образом по характеру биологии питания представляется возможным всех птиц, связанных так или иначе с литоралью, разбить на следующие группы.
I. Птицы, питающиеся в море и на литорали моря плавающими организмами. Птенцы выкармливаются исключительно морскими формами. К ним относится полярная крачка (Sterna paradisea).
II. Птицы, питающиеся на литорали моря, главным образом формами, связанными с грунтом и в меньшей степени свободно плавающими организмами. Птенцы живут на литорали с момента вывода. К этой группе относятся кулики: 1) кулик-сорока (Haemaiopus ostralegus), 2) камнешарка (Arenaria interpres), 3) галстучник (Charadrius hiaticula), 4) малый веретенник (Limosa lapponica) и утки — гага (Somateria mollisima).
III. Птицы, питающиеся в море и на литорали моря свободно плавающими и связанными с грунтом организмами. Наблюдаются довольно регулярные отлеты за пищей в глубь материка. Птенцы выкармливаются как литоральными — морскими, так и не морскими организмами, Сюда относятся чайки: 1) Larus marinus, 2) L. argentatus, 3) L. fuscus и 4) L. canus.
IV. Птицы, питающиеся на литорали моря, главным образом, связанными с грунтом организмами, по прилете (до начала насиживания) и после подъема птенцов на крыло. Но даже и в это время наблюдаются частые полеты за кормом на пресноводные лесные озера. Гнезда устраиваются обычно в лесу на озерах. Птенцы после подъема на крыло живут преимущественно на литорали моря. К ним относятся: кулики — 1) большой улит (Tringa пеbularia) и 2) кулик фи-фи (Т. glareola); утки: 3) турпан (Oidemia fused), 4) чернеть (Nyroca fuligula), 5) гоголь (Clangula clangula) и 6) лебедь-кликун (Суgnus суgnus). С небольшой оговоркой сюда же могут быть отнесены и гагары — 7) Gavia stellata и 8) G. arctica, так как взрослые птицы вылетают на море в течение всего лета, хотя гнезда они строят в большинстве случаев на озерах, где и держатся птенцы до подъема на крыло.
Для всей четвертой группы очень резка изменчивость питания по сезонам.
V. Птицы, частично питающиеся на литорали моря формами, связанными с грунтом. Основное питание — в лесу. Птенцы выкармливаются как лесными, так и литоральными организмами из моря. К этой группе относятся воробьиные: 1) дрозд белобровик (Turdus musicus), 2) зарянка (Erithacus rubecula), 3) серая мухоловка (Muscicapa striata), 4) зяблик (Fringilla coelebs), 5) овсянка (Emberiza citrinella), 6) жулан-сорокопут (Lanius cristatus).
VI. Птицы, питающиеся на литорали моря, но не литоральными формами, а в основном насекомыми. Птенцы выкармливаются исключительно последними. Сюда относится белая трясогузка (Motacilla alba).
Для этого вида литораль моря служит лишь привычным биотопом, похожим на те, где трясогузка обычно добывает себе пищу (песчаные или каменистые берега реки, дороги в сырых местах и другие не заросшие травой и влажные места).
VII. Птицы, питающиеся исключительно в лесу. Наблюдаются лишь спорадические залеты на литораль моря за камешками или растительным кормом (Aster trifolium). Эту группу составляют: 1) белая куропатка (Lagopus lagopus), 2) тетерев (Lyrurus tetrix), 3) глухарь (Tetrao urogalus), 4) рябчик (Tetrastes bonasia).
Методика учета кормов
Наиболее часто применяемым качественным методом учета объектов питания является выяснение количества (в процентах) экземпляров птиц, в желудках которых встретился данный вид пищи. В настоящей работе именно этот метод является основным, хотя он и не дает абсолютно точного соотношения между различными компонентами питания, ибо скорость переваривания и время пребывания в желудке разных видов пищи очень сильно колеблется. В пище многих птиц имеются такие составные части, которые при разборке желудков из-за своей жидкой консистенции вообще не поддаются анализу (яйца), а с другой стороны — такие, которые находятся в желудке очень долгое время и иногда заменяют камни (раковины и т. п.).
Далее необходимо учитывать, что не все то, что находится в желудке, можно рассматривать как пищу. В желудках часто встречается масса случайных продуктов. С этой точки зрения надо, например, рассматривать остатки злаков у чаек.
Анализ питания отдельных видов птиц
I группа
Полярная крачка (Sterna paradisea Brunn). Разобрано 60 желудков из района Беломорской биологической станции (Б. С).
Из данных таблицы 1 видно, что основной пищей крачки является колюшка и нереис.
Н. Горчаковская (1948) указывает, что на Баренцевом море («Семь островов») в рационе крачки существенное место занимает из рыб — песчанка, мойва и сельдь, а из ракообразных — Mysidae.
Таблица 1
Основные компоненты питания полярной крачки (% птиц, в желудках
которых встречен данный вид пищи)
|
Вид пищи |
Встречаемость, % |
|
Рыба (колюшка) |
82 |
|
Нереис (N.pelagica) |
45 |
|
Гаммарус |
17,6 |
|
Насекомые |
11,7 |
|
Всего: виды из моря |
100 |
|
не из моря |
11,7 |
По прилете и до того момента, как колюшка подошла к берегам, крачки питаются, как указывает П. В. Матекин, исключительно гаммарусами (рис. 1), но в дальнейшем их значение в рационе крачки сходит почти на нет. С того момента, как колюшка подходит к берегу для икрометания, крачки питаются в основном ею, но по мере появления в пелагиали гетеронереидной формы нереис, она начинает завоевывать все большее место в рационе этой птицы. К концу лета повышается не только процент частоты встречаемости желудков с нереис, но и среднее количество ее, съедаемое птицей за определенное время.
Следует отметить сравнительно высокий процент питания крачки насекомыми (11.7%).
При сравнении питания молодых и взрослых птиц оказывается, что питание молодых отличается большим однообразием, — в их пище отсутствуют насекомые, а нелетных птенцов родители кормят исключительно колюшкой.
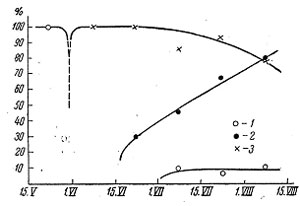
Рис. 1. Сезонные изменения в составе питания крачки – Sterna paradisea. 1 – гаммарус; 2 – нереис; 3 – колюшка.
II группа
1. Кулик-сорока (Haematopus ostralegus ostralegus L.).
Разобрано 19 желудков, из которых десять из района Биостанции и девять из района о-ва Лодейного (общие результаты сведены в табл. 2).
Таблица 2.
Основные компоненты питания кулика-сороки
|
Вид пищи |
Встречаемость, % |
|
Рыба (колюшка) |
5.3 |
|
Нереис |
10.5 |
|
Гаммарус |
15.8 |
|
Моллюски |
100 (во всех желудках) |
|
Насекомые |
31.6 |
|
Растительн. остатки |
36.8 |
|
Всего: виды из моря |
100 |
|
не из моря |
31.6 |
Основным кормом кулика-сороки являются моллюски (примерно в равной степени крупные брюхоногие и двустворчатые) и в меньшей степени насекомые. Свободно плавающие организмы с литорали моря занимают подчиненную роль. Мадон (Madon, 1935), сообщая в общем тот же состав кормов, указывает, что на Северном море этот кулик питается еще крабами и Cardium.
Состав пищи птенцов почти не отличается от пищи взрослых. В ней отсутствуют крупные двустворчатые, которых молодые кулички еще не могут расклевывать.
2. К а м н е ш а р к а (Arenaria interpres interpres L.).
Разобрано 16 желудков: девять из района Биостанции и семь из района о-ва Лодейного (табл. 3).
Таблица 3.
Основные компоненты питания камнешарки
|
Вид пищи |
Встречаемость, % |
|
Рыба (колюшка) |
6.3 |
|
Гаммарус |
37.5 |
|
Моллюски |
87.5 |
|
Насекомые |
31.2 |
|
Раститения |
22.2 |
|
Всего: виды из моря |
100 |
|
не из моря (насекомые) |
31.2 |
Имеются указания, что в других местах, например, на Баренцовом море и на Атлантическом побережье Северной Америки, большое место в рационе камнешарки занимают балянусы. У беломорских камнешарок их встречено не было. Питание взрослых птиц и птенцов ничем существенным не различается.
3. Г а л с т у ч н и к (Charadrius hiaticulah.).
Разобрано 12
желудков: десять из района
Биостанции и два из
района о-ва Лодейного (табл. 4).
Таблица 4.
Основные компоненты питания галстучника
|
Вид пищи |
Встречаемость, % |
|
Рыба (колюшка) |
16.7 |
|
Нереис |
16.7 |
|
Ракообразные |
50 |
|
Моллюски |
91.8 |
|
Насекомые |
70 |
|
Растительные остатки |
18.3 |
|
Всего: виды из моря |
100 |
|
не из моря |
70 |
Этот небольшой куличок предпочитает песчаную или песчано-илистую литораль, изобилующую большим количеством мелких организмов. Из моллюсков он может питаться только мелкими литторинами и гидробиями, которых он съедает вместе со столбиком раковин. Малая питательность этого корма очевидна — он компенсируется по мере встречаемости на литорали различными ракообразными и насекомыми.
4. Малый веретенник (Limosa lapponica lapponica L.).
На Белом море редок. Автором добыт всего один экземпляр, в желудке которого были обнаружены: Jaera albifrons, Macoma baltica и семена водных растений.
5. Гага (Somateria mollisima mollisima L.)f
Разобрано восемь желудков: один из района Биостанции и семь из района о-ва Лодейного (табл. 5).
Таблица 5.
Основные компоненты питания гаги
|
Вид пищи |
Встречаемость, % |
|
Рыба (колюшка) |
12.5 |
|
Гаммарус |
37.5 |
|
Моллюски |
100 |
|
Из них: Мидии |
75 |
|
Литорины (Littorina littorea) |
62.4 |
|
Насекомые |
12.5 |
|
Фукусы |
37.5 |
|
Вороника |
25 |
|
Всего: виды из моря |
100 |
|
не из моря |
37.5 |
Н. Горчаковская (1948) по анализу 29 желудков из заповедника «Семь островов» (Баренцево море) обнаружила примерно ту же закономерность в питании гаги: рыба (песчанка) — 3.45%, мидии — 76%, гаммарус — 3.45%, вороника — 17% и балянус — 31%.
Есть указания, что для баренцевоморских гаг основную роль в питании играют мидии, затем литторины и в меньшей степени гаммарус и балянус.
Следует обратить внимание на довольно частую встречаемость в питании баренцевоморских гаг Balanus balanoidus, в то время как беломорские гаги его совершенно не едят. Причина этого не ясна, но с тем же явлением мы встречаемся у многих птиц (камнешарка, серебристая чайка).
По прилете, пока море не освободится ото льда, гага питается на берегу прошлогодней вороникой. В дальнейшем эта ягода совершенно выпадает из ее рациона.
Сравнивая питание разобранных выше видов птиц II группы, можно видеть, что у них закономерность питания литоральными из моря и не литоральными формами, а также и более мелкими группами кормов примерно одна и та же. Основным кормом являются моллюски (табл. 6), встреченные почти у 100% птиц. Степень питания второстепенными кормами также приблизительно совпадает, хотя у некоторых видов и наблюдаются уклонения от среднего процента для всей группы. Эти уклонения объясняются экологическими особенностями этих видов птиц. Так, например, у галстучника, предпочитающего песчаную, а не каменистую, как другие кулики, литораль, повышается количество съеденных ракообразных и насекомых, а количество моллюсков, наоборот, падает. Это, однако, не меняет общего характера связи галстучника с литоралью, свойственного остальным птицам этой группы.
Таким образом ее можно охарактеризовать как группу, основной пищей которой являются морские моллюски и в меньшей степени ракообразные и насекомые.
Таблица 6.
Основные компоненты питания птиц II группы
|
Вид пищи |
Встречаемость, % |
|
Рыба |
10.2 |
|
Моллюски |
97.3 |
|
Ракообразные |
35.2 |
|
Насекомые |
36.3 |
|
Растительные остатки |
33.9 |
|
Всего: виды из моря |
100 |
|
не из моря (главным образом насекомые) |
38.2 |
III группа
1. Большая чайка (Larus marinus marinus L.).
Данных по питанию этой чайки на Белом море у автора не имеется. Модестов (1939) указывает, что на Баренцовом море, кроме видов пищи, встречающихся у других крупных чаек (L. argentatus, L. fuscas), отмечались частые нахождения мелких птенцов базарных птиц и гаги (до 32% по погадкам). Он наблюдал также, как большая чайка нападает на взрослых моевок. А. Бент (Bent, 1921) также указывает на поедание этой чайкой яиц и птенцов различных птиц.
2. Серебристая чайка (Larus argentatus argentatus Pantopp.).
Разобрано семь желудков: два из района Биостанции и пять из района о-ва Лодейного (табл. 7).
Таблица 7.
Основные компоненты питания серебристой чайки
|
Вид пищи |
Встречаемость, % |
|
Рыба |
85.6 |
|
Моллюски |
57.2 |
|
Звезды |
14.3 |
|
Насекомые |
14.3 |
|
Падаль |
14.3 |
|
Млекопитающие |
14.3 |
|
Всего: виды из моря |
85.6 |
|
не из моря |
43.0 |
Для серебристой чайки района о. Лодейного указывается примерно тот же состав и встречаемость в желудках отдельных кормов. Отмечается также участие в питании крабов. Вообще же набор кормов этой чайки чрезвычайно широк и разнообразен (Модестов, 1939; Bent, 1921; Goethe, 1937). По прилете, пока море еще покрыто льдом, основной ее пищей является вороника. В середине лета ягода полностью выпадает из рациона, но появляется вновь незадолго до отлета. Вообще трудно говорить об облигатности питания серебристой чайки тем или иным кормом. В разные годы и у разных особей оно очень сильно меняется. Так, во время насиживания большую роль в ее рационе играют насекомые (Noll, 1924), которыми в другое время она питается сравнительно мало. В желудках насиживающих птиц также чаще, чем обычно, попадаются крабы.
Что касается питания птенцов, то пуховиков родители кормят исключительно рыбой, в желудках же более взрослых птенцов, кроме рыбы, найдены остатки иглокожих, насекомых и скорлупки яиц.
3. Клуша (Larus fuscus fuscus L.).
Эта чайка сравнительно редко встречается на Белом море. Автором был добыт всего один экземпляр, в желудке которого обнаружены остатки наваги и сельди.
Клуша по общему габитусу и по экологии очень близка к серебристой чайке. Питание ее в общем сходно с подробно разобранным выше питанием серебристой чайки.
4. Сизая чайка (Larus canus canus L.).
Разобрано 16 желудков: восемь из района Ругозерской губы (Биостанция) и восемь из района о-ва Лодейного (табл. 8).
Таблица 8.
Основные компоненты питания сизой чайки
|
Вид пищи |
Встречаемость, % |
|
Рыба |
100 |
|
Нереис |
12.5 |
|
Гаммарус |
12.5 |
|
Моллюски |
18.7 |
|
Насекомые |
25 |
|
Растительные остатки |
56.1 |
|
Вороника |
12.5 |
|
Всего: виды из моря |
100 |
|
не из моря |
37.5 |
Основным компонентом питания этой чайки следует считать рыбу, так же как и у серебристой чайки. Моллюски и насекомые будут уже второстепенными кормами. Птенцы, по данным Н. Горчаковской (1948), выкармливаются мальками рыб, гаммарусами и личинками насекомых.
Довольно трудно говорить о закономерностях питания всей III группы в целом, так как для этого имеется слишком мало материала. Но мне кажется возможным считать, что питание L. fuscus идентично с питанием L. argentatus, на что указывают также Бент (1921) и Коллиндж (Collinge, 1913), а питание L. marinus также сходно с питанием L. argentatus, отличаясь от него только преобладанием крупной рыбы и птенцов птиц (Модестов, 1939).
В среднем для всей III группы можно считать следующий состав пищи и процент желудков, в которых наблюдался тот или другой вид корма (табл. 9).
Таблица 9.
Основные компоненты питания птиц III группы
|
Вид пищи |
Встречаемость, % |
|
Рыба |
90.4 |
|
Моллюски |
41 |
|
Насекомые |
17.9 |
|
Другие не морские компоненты |
23.2 |
|
Всего: виды из моря |
90.4 |
|
не из моря |
41.2 |
Таким образом «основными» кормами этой хорошо очерченной группы являются рыба и моллюски. Набор же «второстепенных» кормов очень широк и сильно колеблется не только по годам и сезонам, но и у разных особей одного и того же вида.
IV группа
Так как имеется очень мало данных по питанию птиц этой группы, то вопрос о степени использования литорали различными видами птиц тотчас по прилете и после подъема молодых на крыло остается открытым, ибо одни виды в это время питаются исключительно на литорали, а другие в какой-то мере используют для кормежки лесные озера, лежащие недалеко от берега.
1. Большой улит (Tringa nebularia Gunn.).
Разобрано два желудка из района Биостанции. В них обнаружены: колюшка, икра колюшки, остатки насекомых, растительные остатки.
2. Кулик фифи (Tringa glareola L.).
Разобрано два желудка из района Биостанции. В них обнаружены: гаммариды, остатки насекомых, растительные остатки.
3. Хохлатая чернеть (Nyroca fuligula L.).
Разобрано пять желудков из района Биостанции. В них обнаружены: прудовики, колюшка, семена и остатки растений.
4. Обыкновенный гоголь (Clangula clangula clangula L.).
Разобрано три желудка из района Биостанции. В них обнаружены: колюшка, остатки насекомых и растительные остатки.
5. Турпан (Oidemia fusca fusca L.).
Разобрано пять желудков из района Биостанции. В них обнаружены: колюшка, прудовики, насекомые и растения.
6. Лебедь-кликун (Суgnus суgnus L.).
Разобрано три желудка молодых лебедей из района оз. Верховского. Все желудки были набиты хвощем.
6. Краснозобая гагара (Gavia stellata Pantopp).
Разобрано три желудка из района Биостанции (один — самки и два — молодых). В желудке самки найдены остатки рыбы. У молодых — остатки насекомых, растения и статолиты крупной рыбы.
Как уже говорилось, гагар можно отнести к этой группе только условно, так как взрослые птицы в течение всего лета регулярно вылетают кормиться на море.
По питанию птиц IV группы на Белом и Баренцовом морях нет более полных данных, настоящих же материалов для суждений о питании всей группы в целом недостаточно. Но во всяком случае общий характер ее связи с литоралью ясен.
Регулярное питание на ней имеет место только ранней весной и осенью, в летнее же время бывают только случайные залеты. Это должно в какой-то мере определить сходный состав кормов, хотя и не такой однородный, как у других групп.
V группа
Эта группа птиц в общей экономике литорали, конечно, играет очень незначительную роль, но интересна тем, что показывает, как легко изменившиеся условия обитания и возможности питания используются птицами.
1. Дрозд-белобровик (Turdus musicus L.).
Разобрано шесть желудков: три взрослых птиц и три молодых из района Биостанции (табл. 10).
Таблица 10.
Основные компоненты питания дрозда-белобровика
|
Вид пищи |
Встречаемость, % |
|
Нереис |
50 |
|
Гаммарус |
50 |
|
Моллюски |
16.7 |
|
Насекомые |
66.6 |
|
Ягоды |
83.5 |
|
Всего: виды из моря |
66.6 |
|
не из моря |
100 |
Такой состав кормов указывает на питание дроздов главным образом насекомыми и ягодами, хотя уже намечается связь с литоралью моря.
2. Зарянка (Erithacus rubecula L.).
Разобран один желудок экземпляра, убитого на литорали моря. Посещение литорали этой птичкой отмечалось не один раз. В желудке обнаружены: остатки насекомых (мухи и жуки) и гаммарус.
3. Обыкновенная овсянка (Emberiza citrinella citronella L.).
Разобрано два желудка из района Биостанции; в них обнаружены насекомые (мухи и жуки), а в одном — две раковинки литторин (L. rudis).
Кроме этих видов птиц на литорали моря случалось наблюдать мухоловку, зяблика, жулана сорокопута, но ни одного экземпляра добыть не удалось.
Таким образом можно сказать, что многие лесные птицы (не говоря уже о таких всеядных, как вороны, для которых одним из основных кормов на Белом море являются мидии), при гнездовании в близком соседстве с морем, довольно широко используют пищевые ресурсы литорали и не только сами питаются на ней, но и выкармливают птенцов литоральными животными.
YI группа
1. Белая трясогузка (Motacilla alba L.)
Разобрано 36 желудков, из которых 31 взрослых птиц и 5 молодых из района Биостанции (табл. 11).
Таблица 11.
Основные компоненты питания белой трясогузки
|
Вид пищи |
Встречаемость, % |
|
Ракообразные |
13.9 |
|
Насекомые |
100 |
|
Растительные остатки |
30.5 |
|
Всего: виды из моря |
13.9 |
|
не из моря |
100 |
Основным кормом трясогузки, как видно из таблицы, являются насекомые. Из литоральных, форм моря встречены только ракообразные, да и то в очень небольших количествах. В желудках птенцов (не летных) были обнаружены только насекомые, а литоральные животные моря не были встречены ни разу. Это указывает на то, что литораль используется трясогузкой только как открытое, не заросшее травой место, удобное для добывания привычной пищи (насекомых), и литоральная фауна как кормовая база совершенно ею не используется.
VII группа'
1. Белая куропатка (Lagopus lagopus L.).
Разобрано три желудка из района Биостанции. В них обнаружены: остатки насекомых, ягоды черники, листья черники и березы, а в одном желудке — Aster trifolium.
Кроме куропаток, автору приходилось видеть на литорали тетерку, а по устному сообщению П. В. Матекина, в районе Биостанции видели на литорали глухаря и рябчиков. Куриные вылетают на литораль моря, чтобы набрать камешков, необходимых им для пищеварения.
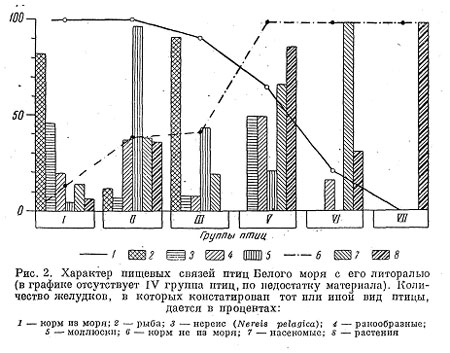
Таким образом, по анализу содержимого желудков и биологической характеристики всех встреченных на литорали Белого моря птиц можно разделить на несколько групп, дающих четкую картину степени связи различных видов с комплексом литоральных животных.
Сравнивая соотношения основных кормов у перечисленных групп птиц, имеющих сходную биологию питания, можно видеть, что у отдельных видов внутри групп оно в общем сходно и в то же время совершенно различно у разных групп. Проведенное разделение по группам позволяет говорить о степени связанности с литоралью уже не отдельных видов, а целых групп птиц, что дает возможность более четко проследить все градации этой связи, не запутанные экологическими особенностями отдельных видов. Можно наблюдать все степени перехода от групп, очень тесно связанных с морем (крачка, кулики и гага), к группам, почти с ним не связанным (рис. 2). Поэтому для Белого моря нельзя провести резкой грани между морской и материковой авиафауной. Нельзя этого сделать и потому, что в рационе «морских» птиц значительное место иногда занимают насекомые, а в рационе типично лесных воробьиных, имеющих возможность использовать литораль моря, большое место занимают морские животные.
Вопросы о смене кормов и использовании в качестве места добывания пищи непривычного биотопа является частью более общего вопроса об устойчивости инстинктов, имеющего в настоящее время большой практический интерес в связи с привлечением птиц в полезащитные лесные насаждения. Полученные данные говорят, что птицы в поисках корма сравнительно легко меняют привычную для них, но бедную кормами стацию, на совершенно новую, но богатую пищевыми ресурсами. Здесь следует отметить, что даже среди таких широко адаптированных птиц, как воробьиные, не все виды могут достаточно легко изменить свои привычки и, в частности, использовать несвойственную им пищу. Причины различного поведения в этом отношении зависят, очевидно, и от различной обеспеченности привычной пищей и от темпов и диапазона приспособляемости к изменению внешних условий, свойственных виду.
Роль птиц в экономике литорали
Большой практический интерес представляет вопрос об удельном весе кормов, съедаемых птицами, по сравнению с их количеством, имеющимся на литорали моря, т. е. вопрос о роли птиц в экономике литорали, а также об относительной интенсивности выедания литоральных животных различными видами птиц.
Для решения вопроса о степени использования этих ресурсов производился количественный учет кормов, съедаемых птицами. Предлагаемый метод подсчета дает возможность судить как об относительной степени интенсивности выедания основных кормов различными видами птиц, так и о роли всего орнитологического комплекса в экономике литорали.
В настоящей работе приводятся лишь ориентировочные количественные данные, так как полному освещению этого вопроса мешают, с одной стороны, скромные количественные данные по питанию, собранные в весьма ограниченный промежуток времени, а с другой — отсутствие экспериментальных данных о скорости переваривания различными птицами разных кормов. Эта величина вообще трудно поддается определению.
У разных видов птиц она различна и изменяется от вида и консистенции пищи, от количества воды, выпитой при ее приеме, и, наконец, от физиологического состояния птицы. Каждое из этих условий может менять скорость пищеварения в несколько раз. Тем не менее, мне кажется возможным дать первое еще весьма приближенное определение значения пищи, съедаемой птицами в общей экономике литорали. Количественные расчеты проводились только для основных литоральных кормов, имеющих к тому же достаточно характерные остатки, по которым можно определить количество животных, съеденных птицей за какой-то определенный промежуток времени. Такой выбор позволяет сравнивать интенсивность выедания литоральных животных различными видами птиц и получить представление о количестве этих кормов, съедаемом за сезон.
Подсчет проводился следующим образом: определялось количество остатков какого-либо корма (например, челюсти нереис или крышечки брюхоногих), приходящееся в среднем на один желудок данного вида птиц. Затем эта величина пересчитывалась на всю популяцию птиц определенного района. Это уже давало возможность сравнивать относительную интенсивность выедания определенного корма разными видами птиц. Зная из литературных данных время пребывания подобной пищи в желудках этого или близких к нему видов, можно было подсчитать количество его, съеденное всей популяцией за сутки и за сезон. Сравнив полученную величину с общей биомассой данного корма на литорали в том же районе, можно говорить о проценте выедания птицами литоральных животных данного вида или об их количестве, выедаемом с единицы площади литорали.
Для выяснения роли птиц в экономике литорали брались наиболее обычные виды в определенном районе Ругозерской губы с площадью зеркала воды 76.5 км2 и с площадью литорали 1.13 км2 (считая ее ширину в среднем равной 25 м).
В качестве примера наших расчетов возьмем литторин, выедаемых популяцией кулика-сороки этого района. В желудке кулика-сороки одновременно находилось в среднем (по 19 желудкам) 2.75 штуки крупных и 4.5 мелких литторин. В рассматриваемом районе питается 15 пар куликов. Скорость опорожнения желудка, по данным Шапошникова (1946) и других авторов, может быть принята равной 8 часам. Таким образом за день кулики съедали 2.75 X 30 х 3=247 крупных и 405 мелких литторин, что при среднем весе L. littorea — 2.07 г, а мелких литторин — 0.08 г составляет 544 г. Время пребывания кулика-сороки на Белом море равняется в среднем 100 дням. Таким образом, можно предположить, что за весь сезон все кулики-сороки этого района съедают 55 кг литторин, т. е. 48.9 кг/км2. По данным Абрикосова и Соколовой (1948), литторины на разбираемом участке дают биомассу в 85.7 г/м2, т. е. 85 700 кг/км2. Следовательно, количество литторин, съеденное куликами за сезон, составляет незначительный процент (0.06%) от общей биомассы. Этот процент выедания моллюсков относится только к взрослым птицам, но даже если принять, что птенцы съедают примерно столько же, хотя это и будет некоторым преувеличением, то и тогда количество съеденных литторин не превысит 0.1% от общей биомассы.
Таким же образом был произведен расчет для других видов птиц (табл. 12).
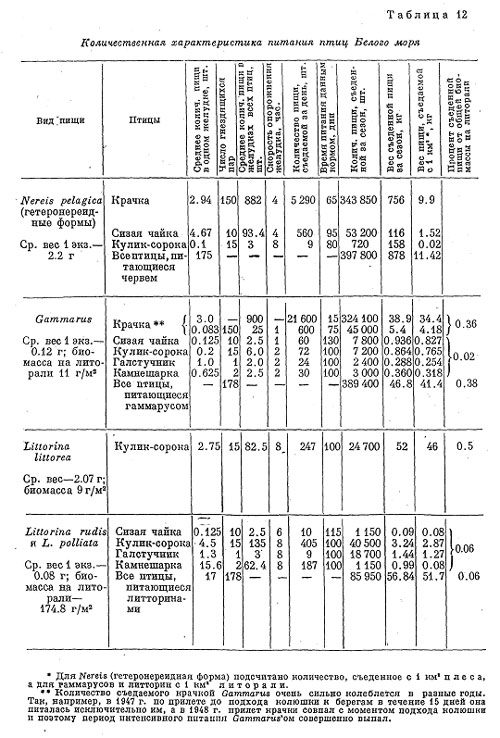
Основными литоральными кормами наиболее обычных видов птиц являются моллюски (брюхоногие и двустворчатые), колюшки, нереис и гаммарус. Отсутствие характерных остатков у колюшки и у двустворчатых делает невозможным их количественный учет при том небольшом материале по питанию их основных потребителей (чайки и гаги), которым мы располагаем. Поэтому в этой таблице имеются данные по выеданию только нереис, гаммарусов и литторин.
Из таблицы видно, что количество литоральных животных, выедаемых птицами, по сравнению со всей их биомассой, очень невелико.
Выводы
1. На основании анализа содержимого желудков и наблюдений по биологии питания оказалось возможным всех гнездящихся в районе Белого моря птиц разделить на несколько групп, отражающих различную степень связи с литоралью моря.
2. Провести резкую границу между морской и материковой орнитофауной Белого моря, основываясь только на питании птиц, оказалось невозможным.
3. Было выяснено, что нередко и лесные птицы используют литоральную фауну в качестве пищевой базы. Вопрос о смене кормов и использовании в качестве места добывания пищи непривычного биотопа является частью более общего вопроса об «устойчивости» инстинктов у птиц.
Полученные данные показывают, что птицы в поисках корма сравнительно легко меняют привычную для них, но бедную пищей стацию на новую, но более богатую пищевыми ресурсами.
4. Для решения вопроса о степени использования птицами пищевых ресурсов литоральной фауны моря производился количественный учет кормов, съедаемых птицами. Предлагаемый в работе метод подсчета дает возможность судить как об относительной степени интенсивности выедания основных кормов различными видами птиц, так и о роли всего орнитологического комплекса в экономике литорали.
5. Полученные результаты позволяют утверждать, что при огромной биомассе литоральных животных, количество их, выедаемое птицами, сравнительно ничтожно —десятые доли процента.
Литература
Абрикосов Г. Г. и С о ко лова Н. Ю. 1948. К изучению фауны литорали Белого моря. Вест. Моск. ун-та, № 2.
Горчаковская Н. 1948. Экология сизой чайки по наблюдениям на Восточном Мурмане («Семь островов»). Охрана природы, № 5.
Гурьянова Е. Ф. 1948. Фауна Белого моря.
Модестов. 1939. Питание чаек восточного Мурмана и роль их в формировании птичьих базаров. Сб. научн. студ. работ МГУ, т. 9.
Шапошников Л. В 1946. К вопросу о ходе опорожнения желудка у птиц. Зоол. журн., т. 26, вып. 4.
Bent А. С. 1921. Life histories of North American gulls and terns. Smitson. Inst., U. S. Nat. Mus., Bull. 113.
Collinge W. 1913. The food of some British wild birds. London.
Goethe Fr. 1937. Beobachtungen und Untersuchungen zur Biologie der Silbermowe (Larus argentatus) auf der Vogelinsel Memmerstand. Journ. fur Ornithologie, Heft 1.
Madоn. 1935. Contribution a l'etude du regime des oiseaux aquatiques. Alauda.
Noll H. 1924. Sumpfvogelleben. Wien — Leipzig — New York.